Suggested Citation: Cherukuri, S. P., Gereau, S., Shostak, L., Hendrickson, S., Frindell, K., & Platkin, C. (2026, May). Defend and heal: Chronic inflammation and cancer [Evidence-based food as medicine research and resource guide]. Center for Food as Medicine & Longevity.
Table of Contents
Introduction
Chronic inflammation is a persistent, low-grade immune response that may persist for months or even years, often in the absence of an apparent trigger. While acute inflammation is a protective and self-limiting response to injury or infection, chronic inflammation is characterized by continuous immune activity that persists without resolution.1 Over time, this dysregulated state contributes significantly to cellular damage and is linked to the development and progression of many major diseases, including cardiovascular disease, diabetes, cancer, and neurodegenerative disorders.2 Recent studies have highlighted the significance of chronic inflammation as a common underlying mechanism in various non-communicable diseases. Of particular concern is its role in cancer development, where it contributes to genetic mutations, tumor growth, and immune system disruption.3 This literature review explores the biological links between chronic inflammation and cancer. It also highlights dietary approaches that can help reduce inflammation and decrease the risk of cancer. By understanding these connections, we can enhance public health initiatives and help individuals make informed lifestyle choices that promote long-term health and prevent disease.
Significance of the Topic
Chronic inflammation is often recognized as a key contributor to the global burden of diseases. It has been identified as a fundamental mechanism underlying many of the world’s most prevalent and deadly diseases with a vast public health impact. According to the WHO, noncommunicable diseases account for over 43 million deaths annually, accounting for approximately 74% of all non-pandemic related deaths globally.4 This alarming figure highlights how prolonged exposure to harmful lifestyle and environmental factors contributes to chronic inflammation and disease burden. Among these deaths, cardiovascular diseases remain the primary cause of mortality. Cardiovascular diseases alone contribute to over 19 million deaths each year, followed closely by cancer.5 In the year 2020, cancer was responsible for nearly 10 million deaths, making it the second leading cause of death globally and accounting for 1 in every 6 deaths.6 More recently, in 2022, there were an estimated 20 million new cases of cancer and a whopping 9.7 million deaths.7 This rising cancer burden has been attributed to aging populations, lifestyle transitions, and increased exposure to pro-inflammatory risk factors. The most common cancer-related deaths are due to lung, colorectal, liver, stomach, and breast cancers. About one-third of all cancer deaths can be prevented by addressing risk factors like tobacco use, alcohol consumption, unhealthy diets, high body mass index, and physical inactivity.8 These modifiable behaviors not only reduce cancer risk but also help regulate inflammatory biomarkers such as C-reactive protein (CRP), IL-6 (Interleukin-6), and TNF-α (Tumor Necrosis Factor – α), which are often elevated in chronic disease states. There is an increasing demand in the scientific community for research to focus on therapies that target inflammation and on integrative methods that tackle the biological pathways behind chronic diseases.
Understanding Chronic Inflammation
At the molecular level, chronic inflammation is characterized by immune dysregulation, where the body’s innate immune response is repeatedly activated. This leads to a continuous secretion of inflammatory markers such as IL-6, TNF-α, and CRP. These markers amplify the inflammatory response and interfere with normal cellular signaling. NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells), a transcription factor that regulates genes involved in immune responses, apoptosis, and cell proliferation, is a major player in this process.9 Consequently, this chronic activation of NF-κB leads to sustained production of cytokines and other inflammatory mediators, resulting in tissue damage. This has made NF-κB a major target for therapeutic interventions, with efforts focused on modulating its activity.10
A variety of modifiable lifestyle and environmental factors significantly contribute to the persistence of chronic inflammation. A few of these include poor diet, physical inactivity, chronic stress, insufficient sleep, smoking, and exposure to environmental toxins such as air pollution and industrial chemicals. A poor diet, especially one high in added sugars, refined carbohydrates, saturated fats, and ultra-processed foods, can promote the production of pro-inflammatory cytokines and disrupt the gut microbiome.11 The trillions of microorganisms in the gut regulate immune function, maintain intestinal barrier integrity, and generate metabolites with anti-inflammatory effects. Disruption of this delicate balance, known as dysbiosis, can initiate or sustain chronic low-grade inflammation.
For instance, added sugars and refined carbohydrates can increase the amount of pro-inflammatory gut bacteria and reduce the amount of anti-inflammatory gut bacteria, which, in turn, contributes to chronic inflammation.12 Similarly, increased consumption of advanced glycation end products (AGEs), which are predominantly found in grilled meats, fried food, and processed snacks, is associated with the development of an inflammatory and pro-oxidative state, potentially leading to the development of chronic diseases.13 A pro-oxidative state refers to a condition in which the balance between oxidants (such as reactive oxygen species, ROS, and reactive nitrogen species, RNS) and antioxidants is shifted toward oxidants, resulting in oxidative stress and chronic inflammation.
On the other hand, a diet rich in anti-inflammatory foods such as fruits, vegetables, whole grains, and healthy fats can help reduce inflammation and decrease the risk of chronic diseases. Collectively, these dietary patterns create an internal environment that promotes inflammation, disrupts metabolic balance, weakens immune regulation, and sets the stage for the development of chronic diseases.
In addition to diet, physical inactivity significantly contributes to the development of low-grade inflammation.14 Sedentary behavior is associated with increased levels of inflammatory markers such as C-reactive protein (CRP), IL-6, and TNF-α, which promote insulin resistance, atherosclerosis, and metabolic dysfunction. Regular physical activity, on the other hand, can help reduce inflammation and improve metabolic health. The increased production of these pro-inflammatory markers has been implicated in the pathogenesis of coronary artery disease (CAD).15 More specifically, elevated levels of CRP have been shown to actively contribute to the pathophysiology of type 2 diabetes by promoting insulin resistance and impairing β-cell function.16
Chronic stress is another major driver of systemic inflammation. Persistent stress activates neuroendocrine pathways, particularly the hypothalamic-pituitary-adrenal (HPA) axis (the body’s central stress response system), leading to elevated cortisol levels and an increased production of reactive oxygen species (ROS).17 This pro-oxidative state can damage cell structures, leading to impaired apoptosis and tumor development. Additionally, stress can also lead to unhealthy coping behaviors such as overeating, poor sleep, and physical inactivity, which further contribute to inflammation and disease risk.
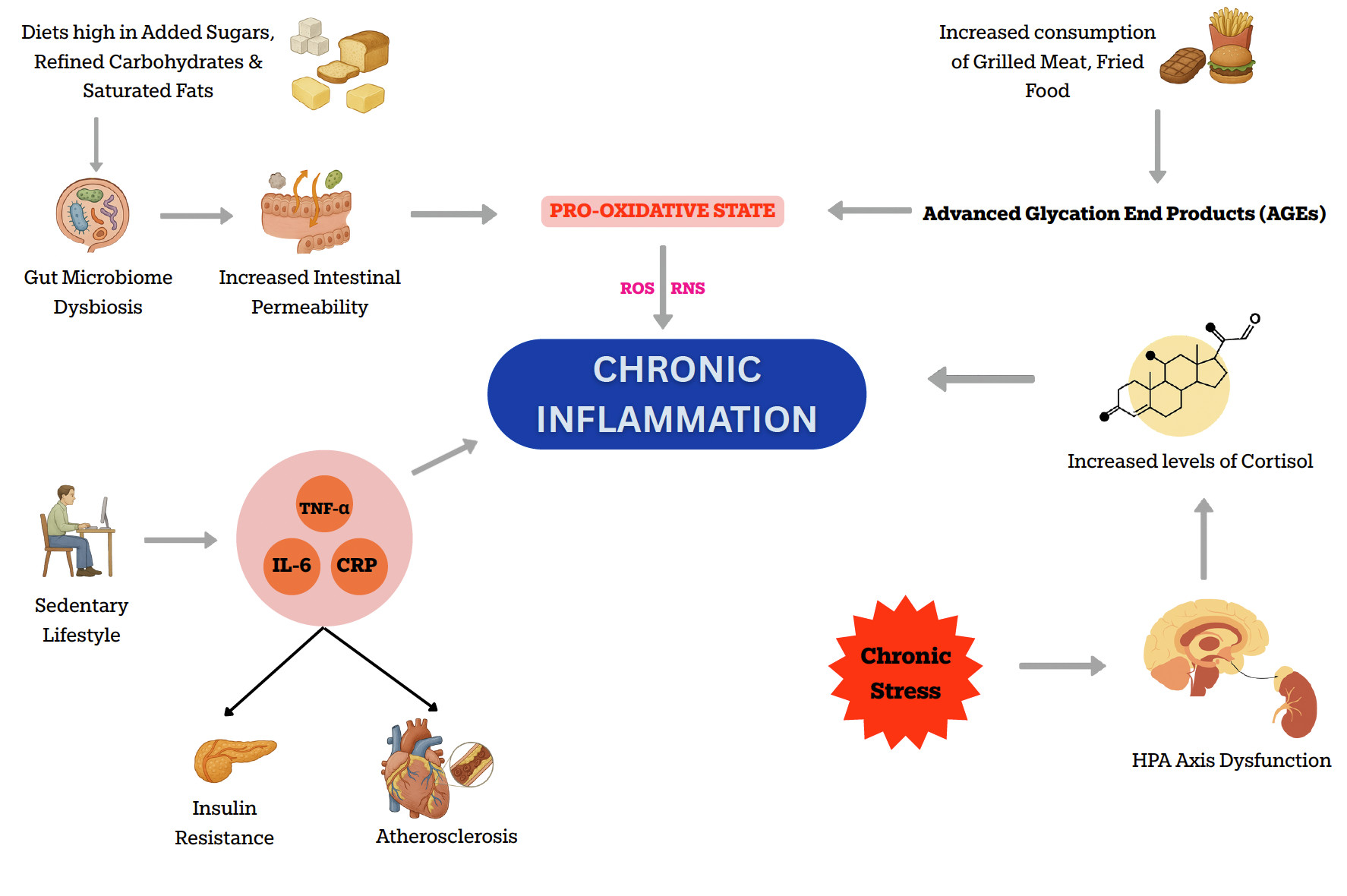
Figure 1: Illustration showing various factors contributing to chronic inflammation
The Link between Inflammation and Cancer
Chronic inflammation plays a critical role in the initiation and progression of cancer.18 Several cancers, mainly those of the colon, breast, lung, and prostate, have well-established links to chronic inflammation.19 Persistent inflammatory stimuli such as infections, autoimmune diseases, or exposure to toxic agents trigger a continuous immune response that disrupts normal tissue homeostasis. Pathogens such as Helicobacter pylori, Epstein-Barr Virus (EBV), and Hepatitis B & C Viruses can contribute to carcinogenesis through mechanisms including chronic inflammation, expression of viral oncogenes, and suppression of host immune responses.20 Autoimmune conditions such as inflammatory bowel disease (IBD), systemic lupus erythematosus (SLE), and rheumatoid arthritis (RA) are well-recognized contributors to chronic inflammation that may promote tumorigenesis in affected tissues.21
Among the toxic agents, tobacco smoke, asbestos, and excessive alcohol consumption have been linked to chronic inflammation and cancer.22 Asbestos exposure primarily occurs in occupational settings, such as the construction and shipbuilding industries. Asbestos fibres are known to induce inflammation and fibrosis in lung tissue, significantly increasing the risk of lung cancers and mesotheliomas, especially when combined with other risk factors like tobacco smoking.23 Crystalline silica is another recognized Group 1 carcinogen (a substance that is definitely carcinogenic to humans) by the IARC (International Agency for Research on Cancer), causing lung cancer through mechanisms involving chronic inflammation and cellular genotoxicity.24 Occupational exposure typically occurs in industries such as mining, construction, sandblasting, and ceramics manufacturing, where inhalation of silica dust is a common occurrence.
Inflammatory processes can change everyday tissue environments into fertile grounds for tumor growth by creating conditions that promote DNA damage, uncontrolled cell proliferation, and impaired apoptosis.25 Pro-inflammatory cytokines such as IL-6, TNF-α, and transcription factors like NF-κB are persistently activated in chronically inflamed tissues.26 Their activation leads to excessive production of reactive oxygen species (ROS) and Reactive Nitrogen Species (RNS) by macrophages and neutrophils.27 These damage the DNA, alter protein structures, and thereby increase the risk of genomic instability (an increased tendency for the cell’s genetic mat and oncogenic mutations. This genomic instability contributes to the accumulation of mutations in oncogenes and tumor suppressor genes.
Mitochondrial dysfunction is a critical intermediary between chronic inflammation and carcinogenesis. Inflammation alters mitochondrial metabolism and leads to the accumulation of ROS, further contributing to oxidative DNA damage, disruption of cellular energy homeostasis, and activation of survival pathways in tumor cells.28 Mitochondria also regulate apoptosis, and their dysregulation promotes increased cell survival and the development of therapy-resistant tumor phenotypes.29 Emerging evidence indicates that inflammation-induced mitochondrial stress not only facilitates tumor initiation but also maintains the inflammatory microenvironment necessary for sustained tumor progression and metastasis.30 Thus, chronic inflammation and mitochondrial dysfunction form a reinforcing feedback loop that amplifies cellular damage and accelerates oncogenic transformation.
Inflammation also supports cellular proliferation by activating signaling pathways, such as STAT3 (Signal Transducer and Activator of Transcription 3) and MAPK (Mitogen-Activated Protein Kinase), key pathways that regulate cell division and growth, that drive cell cycle progression and suppress the activity of tumor suppressor genes. Studies have consistently shown that the MAPK signaling pathway plays a crucial role in cancer progression, treatment response, and resistance.31 Similarly, targeting the STAT3 signaling pathway in cancer immunotherapy holds great promise for enhancing antitumor immune responses and thereby improving the efficacy of cancer treatments.32
Chronic inflammation also promotes the formation of new blood vessels, known as angiogenesis. Angiogenesis plays a crucial role in carcinogenesis by supplying tumor cells with oxygen and nutrients required for growth and metastasis.33 Inhibition of angiogenesis has emerged as a promising therapeutic strategy for improving outcomes in cancer patients. Angiogenesis is mediated by angiogenic factors such as vascular endothelial growth factor (VEGF) and interleukin-8 (IL-8), both of which are upregulated in inflamed and hypoxic environments.34 These molecules increase vascular permeability, allowing growing tumors to receive nutrients and oxygen, thereby further accelerating cancer progression. Additionally, the leaky vasculature created by abnormal angiogenesis can impair drug delivery and facilitate the dissemination of tumor cells into the bloodstream.
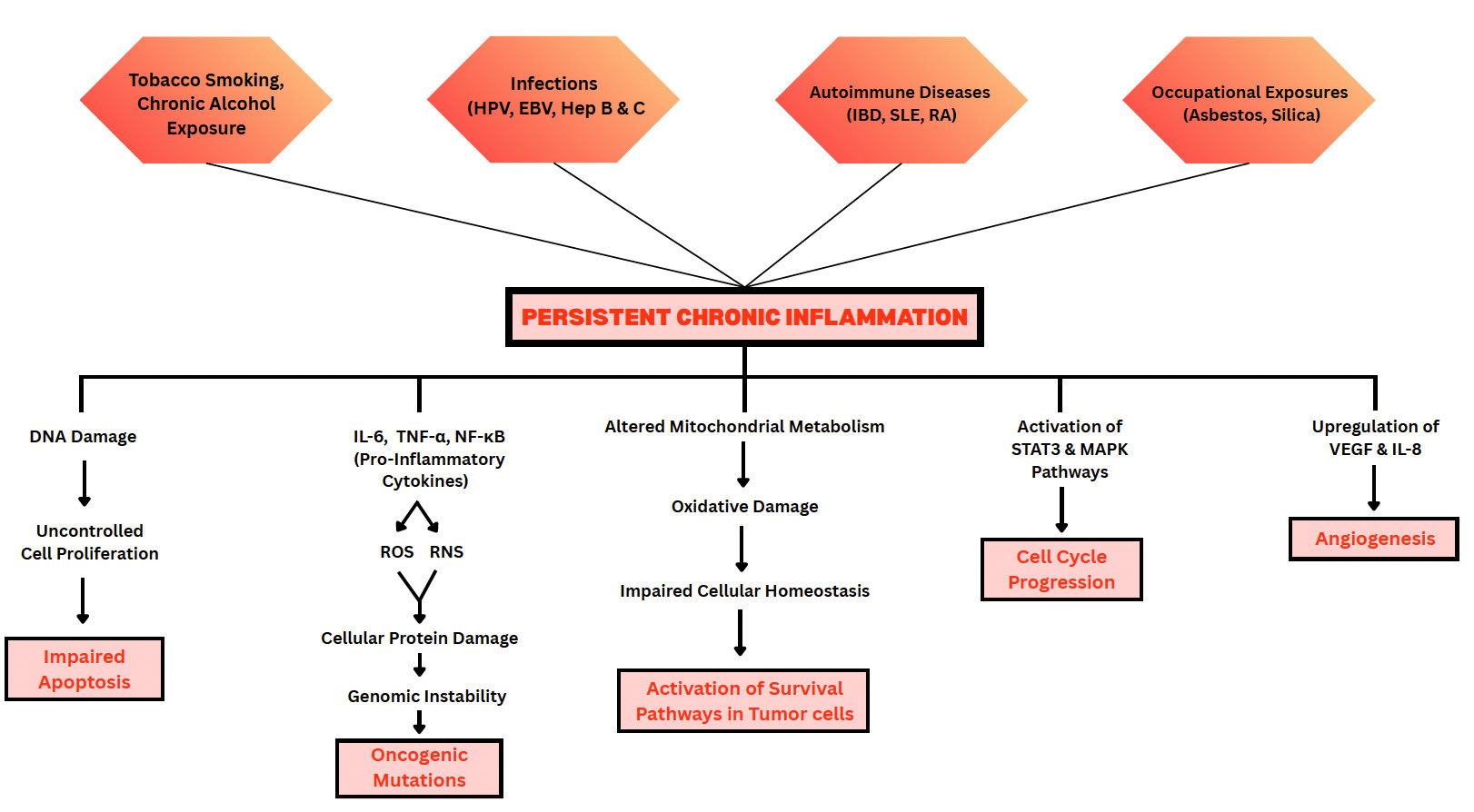
Figure 2: Illustration of how chronic inflammatory stimuli contribute to Oncogenesis.
Specific Nutrient: Curcumin
What is it?
Scientific Name: Curcuma longa L.
Curcumin is the primary active compound found in turmeric, a golden-yellow spice derived from the root of the plant Curcuma longa. Turmeric has been used for over 4,000 years in traditional healing systems, including Ayurveda (from India), Unani medicine (originating from Greece and later developed by Arab and Persian scholars), and Traditional Chinese Medicine.35 These systems have long valued turmeric for its ability to reduce inflammation, support digestive health, aid in skin condition healing, and promote wound healing. In India, turmeric has long been used as a natural remedy for first aid, with people applying its paste to cuts, scrapes, and burns for its antiseptic and anti-inflammatory properties – a practice still common in many communities today.
Beyond its medicinal uses, turmeric is also a beloved kitchen staple in South Asian cooking. It’s warm, earthy flavor and vibrant yellow hue make it an essential in curries, rice dishes, and even herbal teas. Thanks to curcumin, the compound that gives turmeric its color and health benefits, this humble spice continues to gain global attention for its potential in preventing and managing chronic diseases.
How does it work?
Curcumin exerts its effects through a diverse range of molecular pathways that collectively contribute to its antiseptic, antioxidant, anti-inflammatory, and anti-cancer properties. It exhibits broad-spectrum antibacterial effects by disrupting virulence factors, biofilm formation, and microbial growth.36
Curcumin acts as a potent antioxidant through two key mechanisms – direct scavenging of free radicals and upregulation of the body’s antioxidant defenses. Free radicals are usually produced in small amounts during metabolism, but their production increases due to chronic inflammation and stress. The increased production of these molecules can lead to DNA damage, thereby increasing the risk of mutations and cancer. It can also lead to lipid peroxidation, which damages the cell membranes. Excess free radicals also damage protein structures, leading to loss of enzyme function and signaling. Curcumin can directly neutralize reactive oxygen species (ROS), such as the Superoxide anion (O₂⁻) & Hydroxyl radicals (OH), as well as reactive nitrogen species, including Nitric oxide (NO) & Peroxynitrite (ONOO⁻).37 Curcumin also activates genes regulated by Nrf2 (nuclear factor erythroid 2–2-related factor 2), a transcription factor that governs the cellular antioxidant response.38 This dual action – neutralizing free radicals and enhancing internal defenses makes curcumin uniquely effective at protecting cells from oxidative stress and its downstream consequences, such as inflammation and cancer development.
Curcumin is a well-established natural anti-inflammatory agent, primarily due to its ability to modulate critical signaling pathways that regulate inflammation. Its anti-inflammatory and anticancer effects are mediated mainly through the inhibition of the nuclear factor-kappa B (NF-κB) signaling pathway, a central regulator of chronic inflammation and tumor progression. Curcumin blocks the activation of IκB kinase (IKK), thereby preventing the phosphorylation and subsequent degradation of IκBα, which sequesters NF-κB in the cytoplasm. As a result, NF-κB fails to translocate to the nucleus, leading to decreased transcription of pro-inflammatory cytokines, such as TNF-α, IL-1β, and IL-6, as well as pro-inflammatory enzymes like COX-2 and iNOS 39 In addition to inhibiting NF-κB, curcumin modulates other inflammation-related pathways, including MAPK, TLR4, JAK/STAT, and NLRP3 inflammasome activation, further suppressing inflammatory cascades.40
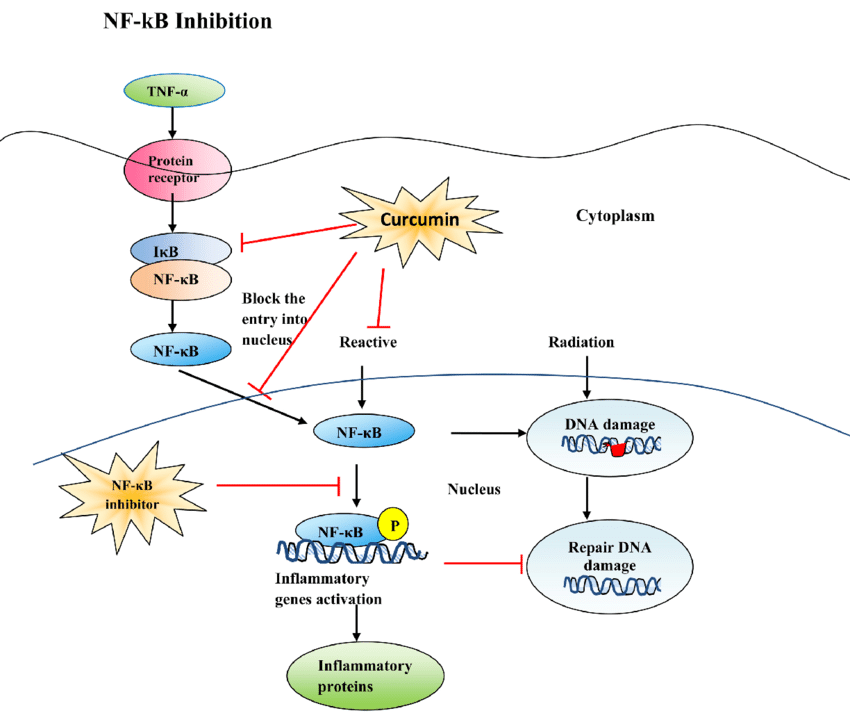
Figure 3: NF-κB Inhibition by Curcumin
Source: https://www.scirp.org/journal/paperinformation?paperid=83874
Beyond inflammation control, curcumin interferes with cancer-promoting processes by downregulating cyclin D1, a key regulator of cell cycle progression, and PECAM-1, a molecule involved in tumor angiogenesis. These actions result in reduced tumor proliferation, diminished micro vessel density, and enhanced apoptosis in cancer cells.41 Curcumin’s ability to act on multiple molecular targets, while maintaining a favorable safety profile, makes it a compelling candidate for both chemoprevention and adjunctive cancer therapy.
What does research tell us?
Curcumin, the active compound in turmeric, has gained attention for its potential role in cancer prevention and treatment. Although curcumin is not a cure, research from animal models and human trials demonstrates its promising effects in slowing tumor growth and reducing precancerous lesions.
A preclinical study conducted in 2015 at Texas Tech University Health Sciences Center investigated curcumin’s impact on triple-negative breast cancer in mice.42 The mice were treated with approximately 25 mg of curcumin per day for six weeks. By the end of the study, mice in the curcumin group showed a statistically significant reduction in tumor volume compared to the control group (p = 0.0195). Histological analysis revealed that curcumin treatment resulted in a decrease in micro vessel density within tumors, indicating a strong anti-angiogenic effect. Molecular testing showed that curcumin suppressed NF-κB activation and downregulated its downstream effectors, including cyclin D1 and PECAM-1, which are associated with tumor proliferation and vascular development. These findings suggest that, at a molecular level, curcumin exerts its anticancer effects by interfering with multiple signaling pathways involved in tumor growth and blood vessel formation.
A Phase I/IIa clinical trial conducted in 2019 at Leicester Royal Infirmary in the UK investigated the safety and potential efficacy of adding curcumin to standard FOLFOX (folinic acid (leucovorin), fluorouracil (5-FU), and oxaliplatin) chemotherapy in patients with unresectable metastatic colorectal cancer.43 The study enrolled 41 patients, with 12 in the initial dose-escalation phase and 29 randomized 2:1 to receive FOLFOX alone or FOLFOX combined with 2 grams of oral curcumin daily. The addition of curcumin was well tolerated and did not result in any increase in serious adverse events. Importantly, patients in the curcumin group experienced improved clinical outcomes, with a median progression-free survival (PFS; the length of time the patient lives without the cancer getting worse) of 291 days compared to 171 days in the control group, and a median overall survival (OS; the length of time the patient is still alive after diagnosis or treatment) increase from 200 days to 502 days. Plasma samples confirmed measurable levels of curcumin, supporting its bioavailability in the chemotherapy setting. This trial is one of the most rigorous demonstrations that curcumin can be safely used in conjunction with chemotherapy and may contribute to improved survival outcomes. Although encouraging, these findings are based on relatively small sample sizes. Larger randomized trials are still needed to confirm curcumin’s role as an adjunct in standard cancer therapy.
In another Phase IIa trial conducted at the University of California, Los Angeles (UCLA) and published in 2011, researchers investigated the effect of oral curcumin in 44 smokers with at least eight aberrant crypt foci (ACF) – early precancerous lesions in the colon.44 Participants received either 2 grams or 4 grams of curcumin capsules daily for 30 days. The study found no significant changes in ACF count in the 2-gram group, but those who received 4 grams per day experienced a 40% reduction in ACF numbers, a statistically significant finding (p < 0.005). Additionally, curcumin was well-tolerated, with no serious adverse effects reported. The researchers also detected curcumin and its metabolites in both plasma and rectal mucosa, indicating systemic and local tissue absorption. This study offers compelling early evidence that curcumin may help reverse precancerous changes in individuals at high risk for colorectal cancer. Despite positive signals, a longer follow-up is needed to determine if reductions in ACF translate to lower cancer incidence.
In a pilot study conducted at the Cleveland Clinic and published in 2006, researchers investigated the effects of curcumin and quercetin supplementation in patients with familial adenomatous polyposis (FAP), a rare hereditary condition characterized by the development of hundreds of precancerous polyps in the colon.45 Five patients were enrolled and received a combination of 480 mg of curcumin and 20 mg of quercetin, taken three times daily for six months. Colonoscopies performed before and after treatment revealed a statistically significant reduction in both the number and size of adenomas. On average, participants experienced a 60% decrease in the number of polyps and a 51% reduction in polyp size (p < 0.05). No serious side effects or adverse lab findings were reported, and compliance was high. Despite its small sample size, the study offered compelling preliminary evidence that curcumin, especially when combined with quercetin, may play a chemopreventive role in genetically high-risk populations. Still, the small sample size highlights the need for larger controlled trials in genetically predisposed individuals.
Lastly, in a multicenter, randomized, double-blind Phase II clinical trial conducted across hospitals in France and published in 2023, researchers evaluated whether curcumin could enhance outcomes in women with advanced or metastatic HER2-negative breast cancer.46 A total of 150 participants were randomized to receive either standard docetaxel chemotherapy with a placebo or with 6 grams of oral curcumin daily. The results were striking: women in the curcumin group had a median overall survival of 21.3 months, compared to 17.4 months in the placebo group – a four-month survival gain that was statistically significant (p < 0.05). Additionally, the objective response rate was higher in the curcumin group (41.5% vs. 26%). Importantly, the supplement was well-tolerated, with no additional toxicity reported. This study is one of the first human trials to demonstrate that curcumin, when used in conjunction with chemotherapy, can significantly extend survival in cancer patients, suggesting a potential role for it as an adjunct in oncology care. Although promising, these findings warrant replication in larger, multi-ethnic cohorts and with more extended follow-up periods.
To better understand curcumin’s overall role in cancer care, a 2023 systematic review evaluated dozens of preclinical and clinical trials involving curcumin in cancer prevention and treatment.47 The review confirmed curcumin’s excellent safety profile, noting its consistent ability to reduce inflammatory markers and oxidative stress. While some trials showed tumor shrinkage, delayed progression, or improved survival, others failed to show significant effects. The authors concluded that curcumin is a promising candidate for adjunctive therapy but emphasized the need for well-powered, placebo-controlled Phase III trials to draw firm conclusions.
One such effort is already underway: a Phase III randomized controlled trial (NCT03769766) is currently evaluating whether curcumin (500 mg twice daily) can delay cancer progression in men with low-risk prostate cancer who are under active surveillance.48 This trial aims to determine whether daily curcumin supplementation can prevent disease progression and may offer some of the most compelling evidence to date on curcumin’s role in cancer chemoprevention.
These studies demonstrate curcumin’s potential as a safe, cost-effective compound with significant anticancer and chemopreventive properties. Evidence from early-phase trials and animal models indicates improvements in tumor control, survival, and a reduction in precancerous lesions. However, larger, multi-center, placebo-controlled studies are essential to confirm these benefits across diverse populations.
Dosage Recommendations Based on Research
Determining an optimal curcumin dosage for therapeutic benefit has been challenging due to its low oral bioavailability.49 Native curcumin is rapidly metabolized in the liver and intestinal wall, leading to limited absorption and minimal systemic availability. Additionally, it undergoes rapid conjugation to glucuronides and sulfates, resulting in low plasma concentrations and a short biological half-life. These pharmacokinetic limitations impact curcumin’s efficacy, even at high oral doses. As a result, standard curcumin preparations often fail to achieve therapeutic levels in target tissues. However, enhanced formulations have improved systemic availability.
Dosing requirements vary by target outcome, ranging from low daily doses for systemic anti-inflammatory support to higher, short-term regimens for cancer prevention or co-therapy. Curcumin doses between 500 and 1,000 mg per day have been shown to significantly reduce systemic inflammation by lowering CRP, TNF-α, IL-6, and malondialdehyde (a common marker of lipid peroxidation and oxidative stress), while improving antioxidant activity, including superoxide dismutase and total antioxidant capacity.50 Randomized controlled trials in patients with osteoarthritis also confirm these effects, with 500 mg twice daily (1,000 mg/day) producing marked improvements in pain, stiffness, and physical function over eight weeks compared to placebo.51 Across studies, this dosage range consistently demonstrates safety and efficacy, supporting 500-1500 mg/day as an effective target for general anti-inflammatory benefit. In individuals with T2DM and metabolic syndrome, Curcumin supplementation at 1,000 mg/day for 8 weeks has been shown to improve lipid profile and glycemic control.52
As a chemopreventive agent, daily doses between 2 and 4 g have shown measurable effects in early clinical studies. Short-term trials in high-risk individuals demonstrated reductions in precancerous colon lesions and adenoma burden, suggesting potential in delaying early tumorigenic changes.53 These findings support the role of curcumin as a safe, low-cost approach for cancer prevention at higher nutritional doses. As an adjunct in chemotherapy, curcumin has been well tolerated at doses of 2–6 g/day when combined with standard regimens such as FOLFOX or docetaxel.54 Clinical studies in colorectal and breast cancers have reported improved treatment response and longer progression-free survival without added toxicity. These results indicate that curcumin may enhance the efficacy and tolerability of conventional therapy while reducing inflammation and oxidative stress associated with chemotherapy.
Table 1: DOSAGE RECOMMENDATIONS BASED ON RESEARCH
| USE/POPULATION | RECOMMENDED INTAKE | DON’T TAKE MORE THAN… | WHEN WILL I SEE RESULTS? |
|---|---|---|---|
| General Anti-Inflammatory Benefit | 500–1,500 mg/day | Up to 2,000 mg/day (unless supervised) | 6–12 weeks for reduction in CRP and improved symptoms |
| Metabolic Syndrome / Type 2 Diabetes | 1,000 mg/day | 2,000 mg/day | 8–12 weeks for improved lipid and glucose profiles |
| Cancer Chemoprevention | 2–4 g/day | 6 g/day (short-term clinical supervision) | 4–8 weeks for biomarker or lesion improvement |
| Adjunct In Chemotherapy | 2–6 g/day (with standard regimens) | 6 g/day | Within treatment cycle; improved tolerance and outcomes |
Facts and Misconceptions
Myth: “Turmeric in food is enough to deliver therapeutic curcumin levels.”
Fact: While turmeric is widely consumed as a spice, its curcumin content is only about 2–5% by weight.55 Achieving therapeutic plasma concentrations used in clinical studies would require consuming tens of grams of turmeric daily, which is impractical and may cause gastrointestinal discomfort. Clinical research consistently demonstrates that supplementation with standardized, bioavailable curcumin formulations is necessary for achieving pharmacological effects.Myth: “Natural equals always safe.”
Fact: Although curcumin is generally well-tolerated, “natural” does not guarantee safety. Curcumin can interact with medications such as anticoagulants (e.g., warfarin), antiplatelets, and certain chemotherapeutic agents.56 Therefore, patients with chronic conditions or those undergoing medical treatments should seek medical supervision before starting high-dose curcumin.Myth: “Curcumin is a cancer cure.”
Fact: Evidence supports curcumin as adjunctive/supportive, not curative. Reviews and trials show pathway modulation and potential to enhance chemo/radiotherapy or reduce side effects, but it does not replace standard cancer treatment.Myth: “All curcumin supplements are equally effective.”
Fact: Bioavailability varies drastically between formulations. Standard curcumin powder is absorbed poorly, whereas curcumin-piperine complexes, liposomal curcumin, nanoparticles, and phytosomes show significantly greater plasma concentrations. Choosing a clinically tested, bioavailable formulation is crucial for efficacy.Myth: “More is always better.”
Fact: Higher doses do not always produce better outcomes and may increase the risk of side effects. Most clinical benefits are achieved with 500–2,000 mg/day of bioavailable curcumin extract. Overdose or long-term intake of curcumin can initiate liver injury through oxidative stress.57
Food Sources
Turmeric (Curcuma longa) is obtained from the underground rhizome (root) of a perennial plant in the ginger family (Zingiberaceae). The rhizomes are harvested, cleaned, boiled or steamed to reduce microbial load and inactivate enzymes, then dried and ground into a fine, golden-yellow powder.58 Despite its rich history as a culinary and medicinal agent, turmeric contains only 2–5% curcumin by weight, making it challenging to achieve therapeutic plasma levels through dietary intake alone.59 Common dietary sources of curcumin include turmeric powder, curry dishes, and golden milk (also known as turmeric lattes), with the average daily intake from food typically providing less than 200 mg of curcumin. Furthermore, curcumin has low water solubility and is poorly absorbed when consumed in isolation. However, its bioavailability can be improved when consumed with dietary fats and piperine, the active component in black pepper, which enhances absorption by inhibiting the metabolism of curcumin. Additionally, traditional curry preparation often involves sautéing turmeric in oil or ghee, which can improve absorption because curcumin is lipophilic; it dissolves in fat, facilitating micelle formation and intestinal uptake.60 Turmeric tea or “golden milk,” made by blending turmeric powder into warm milk or plant-based alternatives, typically provides 60–150 mg of curcumin per cup (from ½–1 tsp turmeric), with lipid content aiding absorption.61
Thus, while turmeric-rich diets may contribute to overall wellness and the control of low-grade inflammation, food-based sources alone are insufficient to achieve the concentrations used in clinical trials targeting chronic diseases or cancer.
Table 2: FOOD SOURCES & THEIR APPROXIMATE CURCUMIN CONTENT
| FOOD SOURCE | APPROXIMATE CURCUMIN CONTENT | NOTES |
|---|---|---|
| Turmeric Powder | 2-5% by weight | Most common dietary source; curcumin is poorly absorbed unless taken with fat or piperine |
| Curry dishes (with turmeric) | Varies; often <100 mg/serving | Bioavailability depends on the fat and spice blend used |
| Turmeric tea (“golden milk”) | 60–100 mg per serving | Improved when consumed with milk or coconut oil |
Supplemental Sources
Curcumin supplements are available in multiple delivery forms, each designed to address the inherently low bioavailability. Pharmacological approaches, particularly nano formulations, have been shown to significantly enhance the oral bioavailability of curcumin, thereby improving its therapeutic potential and enabling effective coadministration with complementary adjuvants.62 Nanoparticle-based formulations, such as Theracurmin, which embed curcumin in colloidal submicron particles, have demonstrated approximately 27-fold higher systemic availability and significantly elevated plasma concentrations without added toxicity in Phase I cancer trials.63 Similarly, a seminal study in healthy volunteers demonstrated that co-administering 2g of curcumin along with 20mg of piperine (black pepper extract) increased oral bioavailability by approximately 2000% compared to curcumin alone.64
Another widely adopted strategy is the use of curcumin-phosphatidylcholine complexes, such as Meriva.65 These phytosomal formulations have shown a ~29-fold increase in absorption compared to unformulated curcumin in human pharmacokinetic studies.66 Additionally, liquid micellar and liposomal formulations have also significantly increased curcumin absorption. In a healthy volunteer crossover study, micellar formulations provided several-fold greater bioavailability versus micronized powder.67 A 2023 review further reinforces these findings, highlighting nano formulations as the most promising strategy for enhancing the clinical efficacy of curcumin in cancer therapeutics.68
Table 3: COMPARATIVE SUMMARY OF CURCUMIN FORMULATIONS
| FORMULATION TYPE | MECHANISM / TECHNOLOGY | BIOAVAILABILITY IMPROVEMENT |
CLINICAL EVIDENCE |
|---|---|---|---|
| Theracurmin | Nanoparticle, colloidal submicron particles | ~27× systemic availability | Phase I trial in cancer patients – elevated plasma levels, no added toxicity |
| Curcumin + Piperine | Bioenhancer (black pepper extract) inhibits metabolism | ~2000% increase in healthy volunteers | Human study with 2g curcumin + 20mg piperine |
| Meriva (Phytosome Complex) | Curcumin bound to phosphatidylcholine | ~29× absorption improvement | Randomized double-blind pharmacokinetic studies in humans |
| Micellar Formulations | Solubilized in micelles for better absorption | Several-fold increase vs. micronized powder | Crossover study in healthy volunteers |
Side Effects
Curcumin is generally considered safe and well-tolerated in both healthy individuals and patients, even at high doses up to 6–8 grams per day in short-term studies. However, mild gastrointestinal symptoms such as bloating, diarrhea, and nausea have been reported, particularly at doses above 4 grams daily.69 Some individuals may also notice harmless yellow discoloration of stool due to curcumin’s pigment. Importantly, curcumin may interact with anticoagulant or antiplatelet drugs, increasing the risk of bleeding, and should be used with caution in individuals with gallbladder disease or those undergoing surgery. Curcumin can interact with anticoagulants by both pharmacodynamic (additive anticoagulant effect) and pharmacokinetic (altering drug metabolism via cytochrome P450 inhibition) mechanisms.70 This can lead to unpredictable increases in anticoagulant activity and bleeding risk, especially with drugs that have a narrow therapeutic window. While no serious toxicities have been consistently reported, clinical supervision is recommended when curcumin is used alongside chemotherapy or in high-risk populations.
Table 4: COMMON ADVERSE EFFECTS OF CURCUMIN
| ADVERSE EFFECTS | RISK LEVEL | REPORTED AT |
|---|---|---|
| Mild GI symptoms (nausea, bloating) | Low | ≥ 4,000 mg/day |
| Interaction with anticoagulants | Moderate | Co-administration |
| Risk in gallbladder disease | Moderate | May increase bile secretion |
Legal Status and Availability
Curcumin is legally classified as a dietary supplement rather than a pharmaceutical drug in many countries, including the United States, Canada, Australia, and European Union nations. In the U.S., the FDA has designated curcumin as “Generally Recognized as Safe” (GRAS) when used in food products.71 It is commonly sold over the counter in various forms, including capsules, tablets, powders, and functional beverages. However, curcumin supplements are not FDA-approved for the treatment of any disease, and their quality and potency may vary significantly between manufacturers due to limited regulation under the Dietary Supplement Health and Education Act (DSHEA) of 1994.72
Bottom Line
Curcumin is a safe and cost-effective compound with strong preclinical evidence and encouraging early clinical data, demonstrating anti-inflammatory, antioxidant, and anticancer properties. While it cannot replace standard therapies, studies suggest that it may reduce precancerous lesions, improve progression-free and overall survival when combined with chemotherapy, and lower systemic inflammation. However, its clinical use is still limited by poor oral bioavailability, variable supplement quality, and the need for more large-scale randomized trials. For now, curcumin should be viewed as a complementary strategy, best utilized in bioavailable formulations and under medical supervision, to support, but not replace, standard cancer prevention and treatment approaches.
Specific Nutrient: Omega-3 Fatty Acids
What is it?
Omega-3 polyunsaturated fatty acids (PUFAs), commonly known as Omega-3 Fatty Acids or Omega-3s, are essential nutrients primarily found in marine and certain plant sources. Omega-3s are essential fatty acids, meaning they cannot be synthesized by the human body and must be obtained from the diet. The three main types of omega-3 fatty acids are: Alpha-linolenic acid (ALA), Eicosapentaenoic acid (EPA), and Docosahexaenoic acid (DHA).73 ALA is primarily sourced from plant-based foods, notably flaxseed, chia seeds, hempseed, and walnuts.74 EPA and DHA are concentrated in marine sources, including fatty fish like salmon, trout, and mackerel, as well as certain types of algae.75 Although the human body can convert ALA into EPA and DHA, this conversion is highly inefficient, with rates generally well below 15%.76 As a result, relying solely on plant-based sources of ALA often does not provide adequate levels of EPA and DHA to meet physiological demands, particularly for cardiovascular, neurological, and visual health. For this reason, many individuals, especially those with limited fish or seafood intake, may benefit from omega-3 supplementation (e.g., fish oil or algal oil), since obtaining enough from whole foods alone can be challenging.
Figure 4: Essential Fatty Acids and Dietary Sources.
Source: https://lpi.oregonstate.edu/mic/other-nutrients/essential-fatty-acids
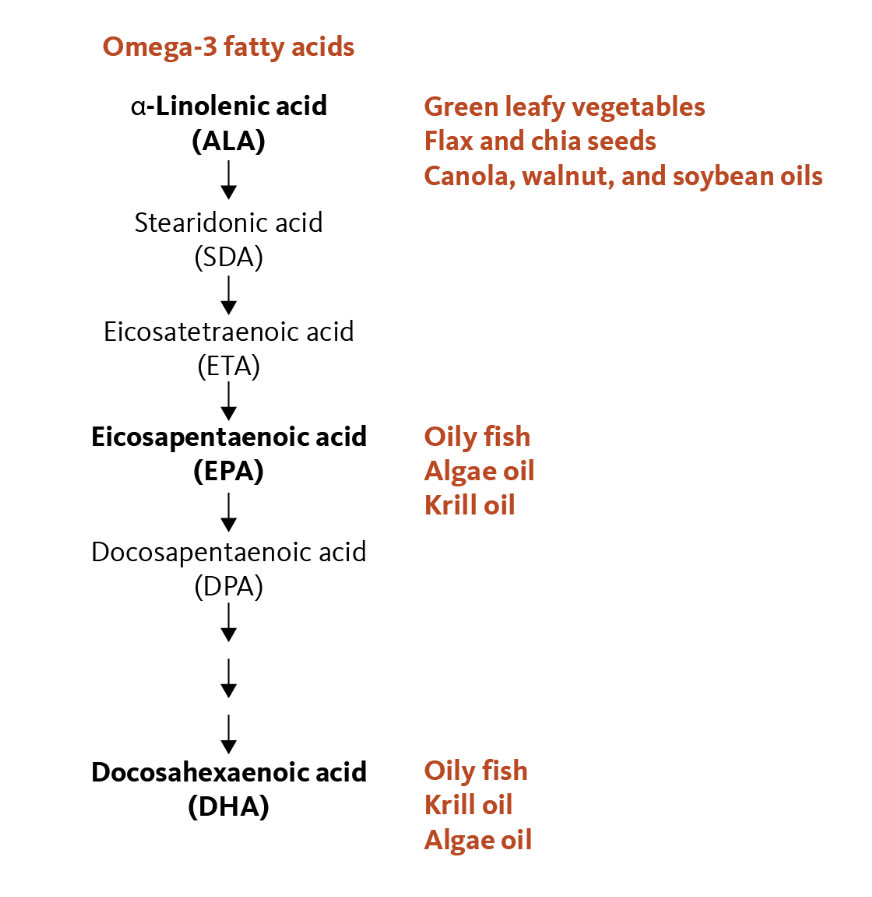
For centuries, cultures with high fish consumption, from the Inuit of Greenland to coastal Japanese populations, have benefited from omega–3–rich diets, which have been historically associated with a reduced risk of cardiovascular and inflammatory diseases. Although traditional diets featuring fatty fish, walnuts, flaxseeds, and chia have long provided these nutrients, it is only in recent decades that scientists have begun to uncover their molecular role in reducing inflammation, supporting brain health, and modulating immune responses. Today, omega-3 fatty acids are not only recognized as vital nutrients for general health but are also being investigated for their potential in preventing and managing chronic diseases, including cancer.
How does it work?
Omega-3 fatty acids exert their effects at both the cellular and molecular levels. Once incorporated into cell membranes, particularly in immune and endothelial cells (the cells lining blood vessels), they influence membrane fluidity, a change that enhances cellular function and signaling.77 EPA and DHA serve as substrates for the production of specialized lipid mediators such as resolvins, protectins, and maresins (signaling molecules derived from omega-3s that actively promote the termination of inflammation) that actively resolve inflammation rather than simply suppress it.78 This pro-resolving activity helps terminate chronic inflammatory responses that, if unchecked, can contribute to tissue damage and cancer development.
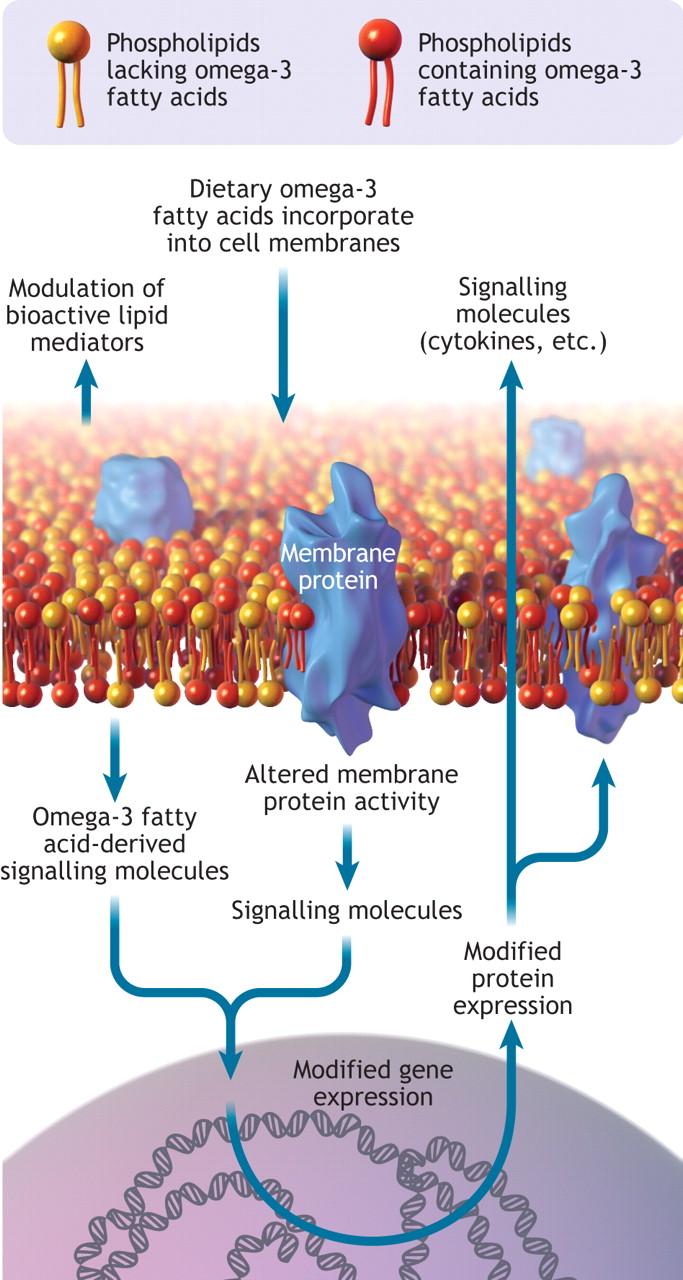
Figure 5: Cell membrane showing omega-3 fatty acids incorporated into the phospholipid bilayer. Omega-3 fatty acids can modify gene and protein expression, modulate membrane protein activity and act as a reservoir for bioactive molecules.
Source: https://www.cmaj.ca/content/178/2/177
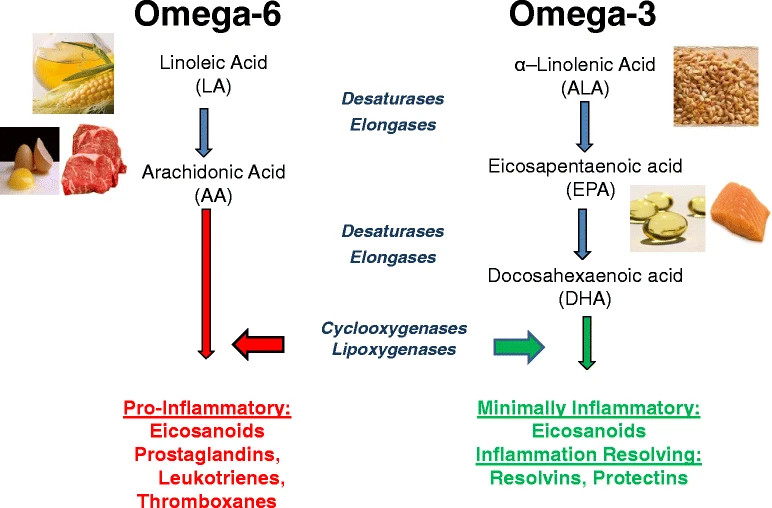
Alongside omega-3 fatty acids, omega-6 polyunsaturated fatty acids (PUFAs) constitute the other major class of essential fatty acids required for human health. They are primarily derived from plant oils such as soybean, corn, safflower, and sunflower oil, and are vital for cell membrane structure, growth, and immune function.79 However, excessive consumption of omega-6s, particularly linoleic acid, can favor the production of pro-inflammatory lipid mediators when not balanced by adequate omega-3 intake.80 Epidemiological and mechanistic studies show that a high dietary omega-6 : omega-3 ratio, often exceeding 15:1 in Western diets, correlates with increased inflammatory activity and higher risk of chronic diseases, whereas a more balanced ratio closer to 4:1 supports optimal immune and vascular function.81
In addition, omega-3s compete with arachidonic acid (an omega-6 fatty acid) for cyclooxygenase (COX) and lipoxygenase (LOX) enzymes.82 While arachidonic acid metabolism yields pro-inflammatory eicosanoids (like prostaglandin E2 and leukotriene B4), omega-3 metabolism produces less inflammatory counterparts, shifting the balance toward an anti-inflammatory state. Beyond inflammation, DHA is integral to neuronal membranes, supporting synaptic plasticity and cognitive function.83 EPA contributes to cardiovascular health by lowering triglycerides, improving endothelial function, and reducing platelet aggregation.84 Together, these mechanisms highlight omega-3 fatty acids as critical mediators of both health maintenance and disease prevention.
Figure 6: Dietary sources and general metabolic pathway for omega-6 and omega-3 polyunsaturated fatty acids, leading to proinflammatory and anti-inflammatory products, respectively.
Source: https://link.springer.com/article/10.1186/s13058-015-0571-6
In addition to their role in resolving inflammation, omega-3 fatty acids exhibit several mechanisms with potential anti-cancer activity. By incorporating into tumor cell membranes, EPA and DHA alter membrane architecture, thereby disrupting pathways critical for cancer cell proliferation and survival. At the same time, they induce apoptosis through mitochondrial pathways: increasing reactive oxygen species (ROS), activating caspases (a family of enzymes that play a role in inflammation and apoptosis), and reducing anti-apoptotic proteins like Bcl-2 and survivin. These combined effects shift the balance toward programmed cell death and limit uncontrolled tumor growth.
Omega-3s also inhibit angiogenesis by downregulating vascular endothelial growth factor (VEGF), platelet-derived growth factor (PDGF), and matrix metalloproteinases (MMPs), which are essential for new blood vessel formation and extracellular matrix remodeling. At the genetic level, omega-3s downregulate nuclear factor-kappa B (NF-κB), a transcription factor that controls the expression of many pro-inflammatory cytokines, chemokines, and adhesion molecules.85 Similarly, inhibition of STAT3 signaling reduces expression of proliferative and angiogenic mediators.
By modulating NF-κB and STAT-3 pathways, omega-3s help reduce systemic inflammation, regulate immune responses, and create a cellular environment less favorable for tumor initiation and progression. EPA and DHA are precursors for bioactive lipid mediators such as resolvins and protectins, which actively resolve inflammation. In the tumor context, these mediators reduce infiltration of immunosuppressive M2 macrophages and myeloid-derived suppressor cells (MDSCs), while promoting cytotoxic T-cell activity and natural killer (NK) cell responses.
Emerging evidence indicates that omega-3 fatty acids can enhance the efficacy of standard chemotherapy while reducing treatment-related toxicity. DHA, for example, increases the sensitivity of cancer cells to drugs like doxorubicin and cisplatin by disrupting cell membrane structure and drug efflux pumps, thereby improving intracellular drug accumulation. Omega-3s also help protect normal tissues by reducing chemotherapy-induced inflammation, cachexia, and neurotoxicity. This dual role, sensitizing tumor cells while protecting healthy cells, makes omega-3s a promising adjuvant in cancer therapy.
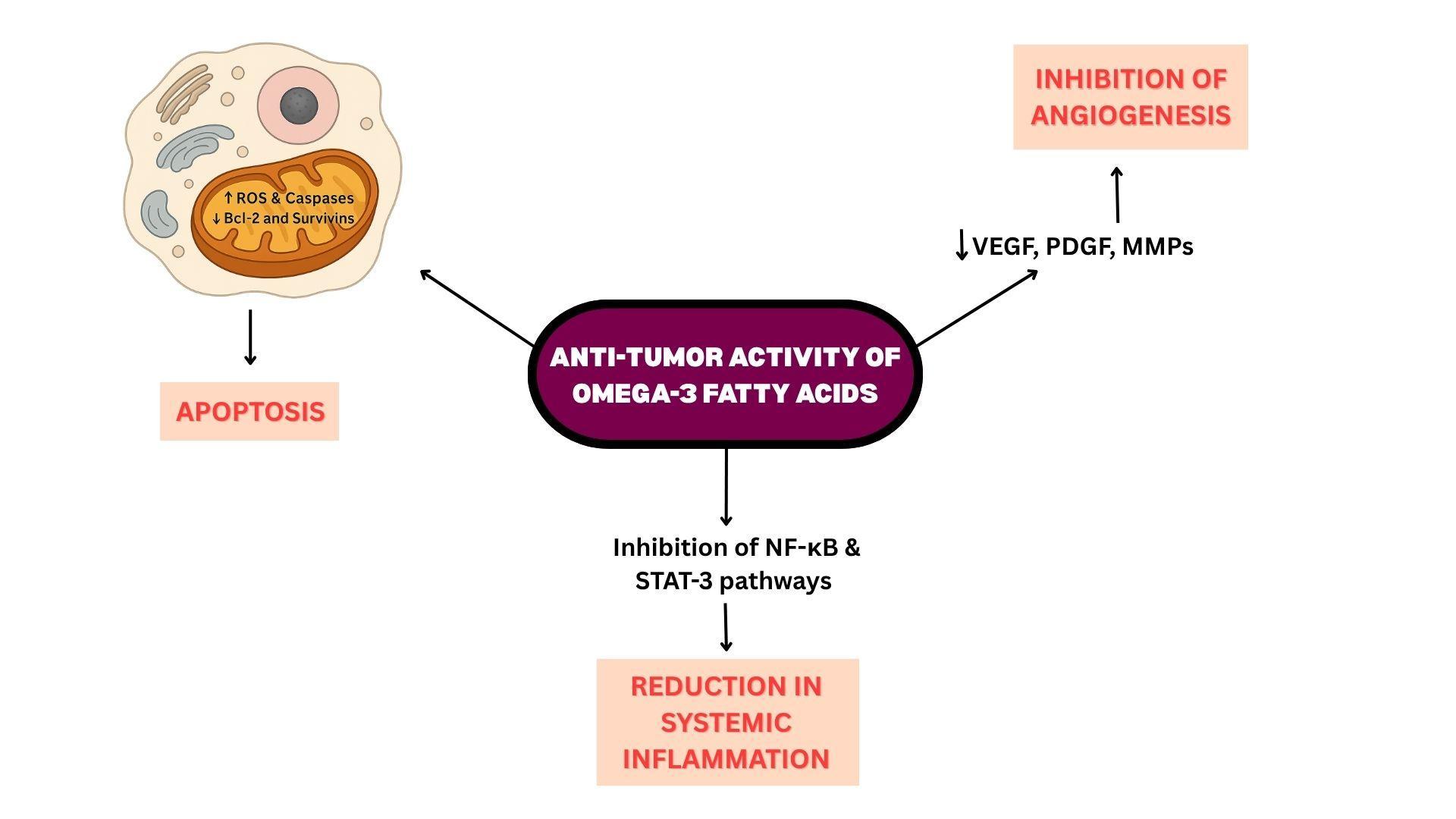
Figure 7: Anti-Tumor Activity of Omega-3 Fatty Acids
What does research tell us?
Omega-3 fatty acids have become the focus of numerous large-scale cohort studies and targeted oncology trials aimed at understanding their potential in cancer prevention and progression. The NIH–AARP Diet and Health Study, one of the largest U.S.-based cohorts with over half a million participants, has provided strong epidemiological evidence.86 The NIH–AARP Diet and Health Study is a prospective cohort study undertaken to investigate diet, lifestyle, and cancer outcomes. Initiated between 1995 and 1996 by the National Cancer Institute in collaboration with AARP, the study enrolled 566,402 men and women aged 50–71 years from six U.S. states (California, Florida, Louisiana, New Jersey, North Carolina, Pennsylvania) and two metropolitan areas (Atlanta and Detroit). Participants completed detailed food-frequency and lifestyle questionnaires at baseline, and cancer outcomes were tracked longitudinally through state cancer registries and the National Death Index, providing robust follow-up data extending more than two decades.
The overarching focus of the NIH–AARP study was to clarify the relationship between dietary patterns, specific nutrients, and cancer risk across multiple organ systems. Outcomes evaluated included incidence and mortality for colorectal, breast, prostate, pancreatic, and liver cancers, as well as mortality from chronic liver disease. Within this large framework, fish consumption and long-chain omega-3 fatty acid intake were studied extensively as exposures of interest. Analyses of 293,464 men in the cohort revealed that higher consumption of fish and long-chain omega-3 fatty acids was associated with a lower risk of fatal prostate cancer, though no consistent association was observed with overall prostate cancer incidence.87 These findings suggest that omega-3s may not prevent the initial development of prostate cancer but could attenuate its progression to lethal forms.
The Singapore Chinese Health Study (SCHS) is another prospective cohort designed to investigate diet, lifestyle, and cancer risk in an Asian population.88 Between 1993 and 1998, the study enrolled 63,257 Chinese men and women aged 45–74 years from the Hokkien and Cantonese dialect groups residing in Singapore. Dietary intake was assessed using a culturally tailored, validated 165-item food-frequency questionnaire, and incident cancer cases were identified through the Singapore Cancer Registry, enabling long-term follow-up. Among 35,298 women included in an early analysis of breast cancer risk, those in the top three quartiles of long-chain omega-3 fatty acid intake had a 26% lower risk of developing breast cancer compared with women in the lowest quartile, after an average of 5.3 years of follow-up.89 The protective association was strongest among postmenopausal women, while higher intakes of animal fat were associated with increased risk. These findings provided some of the first prospective evidence from an Asian population that consuming omega-3-rich fish may reduce the risk of breast cancer.
The VITamins And Lifestyle (VITAL) Cohort, established in Washington State between 2000 and 2002, enrolled more than 77,000 adults aged 50–76 years to investigate how long-term supplement use influences cancer risk.90 Participants completed a detailed baseline questionnaire that captured their use of vitamins, minerals, and specialty supplements, including fish oil, as well as their diet, lifestyle, and medical history. Supplement users were targeted in recruitment: 66% used multivitamins, 46% used individual vitamin C, 47% used individual vitamin E, and 46% used calcium, typically for 5–8 of the past 10 years. Cancer outcomes were ascertained through linkage with the regional SEER cancer registry, providing reliable prospective follow-up.
Analyses from this cohort revealed notable associations with omega-3 fatty acids. In an analysis of 35,016 postmenopausal women, current fish oil supplement use was associated with a 32% lower incidence of invasive breast cancer, with the strongest reduction seen for ductal carcinoma.91 Dietary omega-3 intake from fish, however, was not significantly associated with decreased risk, suggesting that supplement exposure may have been more relevant in this population.
This discrepancy may reflect differences in exposure measurement as fish intake estimates from food-frequency questionnaires are prone to misclassification, whereas supplement use provides a more consistent and quantifiable source of omega-3s. It is also possible that supplement users achieved higher or more sustained intake levels than were typically obtained through diet alone, or that supplement use correlated with other health-conscious behaviors that influenced cancer risk.
Additional work from the VITAL Cohort also indicated that higher intake of long-chain omega-3 was linked with a modest reduction in colorectal cancer risk. This finding was particularly notable among individuals with lower genetic risk scores.92 While observational in nature and unable to prove causality, the VITAL Cohort contributed to the growing body of evidence suggesting that omega-3 fatty acids, particularly through fish oil supplementation, may play a role in reducing the risk of both breast and colorectal cancers.
A noted RCT is the Vitamin D and Omega-3 Trial (VITAL), conducted in the United States between 2010 and 2018, which was the largest randomized, double-blind, placebo-controlled trial to date evaluating whether marine omega-3 fatty acids could prevent cancer in generally healthy adults.93 The study enrolled 25,871 men aged 50 years and older and women aged 55 years and older, who were randomized to receive daily omega-3 supplements (1 g EPA+DHA), vitamin D3 (2,000 IU), both agents, or placebo, with a median follow-up of 5.3 years. Omega-3 supplementation did not significantly reduce the incidence of total invasive cancer, nor was there evidence of protection against breast, prostate, or colorectal cancer specifically. A modest, nonsignificant trend toward lower cancer mortality was observed after excluding early deaths, raising the possibility that omega-3s may play a greater role in slowing cancer progression than in preventing tumorigenesis.
Another prominent RCT is the DO-HEALTH trial (Vitamin D3–Omega-3–Home Exercise–Healthy Ageing and Longevity Trial) is a multinational, randomized, double-blind, placebo-controlled study of 2,157 community-dwelling adults aged ≥70 years recruited from seven European cities – Zürich, Basel, Geneva, Toulouse, Berlin, Innsbruck, and Coimbra.94 Participants were assigned in a 2×2×2 factorial design to vitamin D3 (2,000 IU/day), omega-3 fatty acids (1 g/day; EPA: DHA 1:2 from marine algae), and/or a simple home exercise program (strength training 30 minutes, 3×/week) for three years. The five primary endpoints were incident non-vertebral fractures, functional decline (SPPB), change in blood pressure, cognitive decline (MoCA), and infections; cancer was evaluated as a pre-defined exploratory outcome. In that exploratory analysis, the combination of vitamin D3 + omega-3 + exercise produced a 61% reduction in the risk of any verified invasive cancer versus placebo, while each intervention alone showed only small individual benefits.
Building on evidence from large observational cohorts and RCTs, more recent clinical trials have begun to test the role of omega-3 fatty acids in cancer progression and prevention. The CAPFISH-3 trial, a randomized clinical study of men with low-risk prostate cancer on active surveillance, investigated whether a high omega-3, low omega-6 diet supplemented with fish oil could alter tumor biology.95 After one year of intervention, men in the omega-3 arm showed a significant reduction in Ki-67, a tumor proliferation marker strongly associated with disease progression and mortality.
In parallel, the COLYNE trial is testing the preventive potential of omega-3s in individuals with Lynch syndrome, a hereditary condition that confers high lifetime risk of colorectal cancer. This ongoing randomized study is evaluating whether daily omega-3 ethyl ester supplementation can reduce adenoma formation or delay cancer onset in genetically predisposed patients.96 Although results are still pending, the trial is designed to provide critical insight into whether omega-3s can be deployed as a safe, low-cost strategy for colorectal cancer prevention in high-risk populations.
Some study findings have suggested omega-3s might increase cancer risk – mostly based on biomarker studies in prostate cancer. Two often-cited analyses found higher circulating long-chain omega-3s associated with higher prostate cancer risk: a PCPT (Prostate Cancer Prevention Trial) nested case-control study (DHA and high-grade disease) and the SELECT case-cohort analysis (higher plasma EPA/DPA/DHA and total/high-grade disease).97 98 Although analyses from the PCPT and the SELECT trial suggested that higher long-chain omega-3 levels might be linked to increased prostate cancer risk, many scientists have questioned the significance of these findings.
In the SELECT analysis, for example, the difference in omega-3 levels between men with and without prostate cancer was very small and likely of limited physiological relevance. Others have noted that localized prostate cancers, even high-grade tumors, often progress slowly and are frequently detected at autopsy in men who died of unrelated causes, making mortality a more meaningful endpoint than incidence. In addition, desaturation enzymes that convert ALA to EPA and DHA can be upregulated in cancer cells, raising the possibility that the disease itself may increase omega-3 levels, rather than omega-3s driving disease risk.
Dosage Recommendations Based on Research
The optimal intake of omega-3 fatty acids depends on physiological needs and intended health outcomes. The National Institutes of Health (NIH) Office of Dietary Supplements emphasizes that the United States has no official Dietary Reference Intakes (DRIs) for the long-chain omega-3 fatty acids eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA).99 Instead, the only established Adequate Intakes (AIs) apply to α-linolenic acid (ALA) – 1.6 g/day for men and 1.1 g/day for women which serves as the essential dietary precursor for EPA and DHA. Because endogenous conversion of ALA to long-chain omega-3s is inefficient, international authorities recommend obtaining pre-formed EPA and DHA from fish or algal sources. The World Health Organization/Food and Agriculture Organization (WHO/FAO) Expert Consultation on Fats and Fatty Acids in Human Nutrition specifies that adult males and non-pregnant, non-lactating females should consume at least 0.25 g/day (250 mg/day) of combined EPA + DHA to reduce coronary heart-disease risk, and identifies an acceptable macronutrient distribution range (AMDR) up to 2 g/day, noting that intakes as high as 3 g/day have not been associated with adverse effects.100 The European Food Safety Authority (EFSA) similarly recommends 250 mg/day EPA + DHA for adults to maintain normal cardiac function.101 The American Heart Association (AHA) advises two servings of fatty fish per week (≈ 500 mg/day EPA + DHA) for adults without coronary disease and 1 g/day for those with established coronary heart disease.102
Table 5: ADEQUATE INTAKES (AIS) FOR OMEGA-3S (ALA) 103
| AGE | MALE | FEMALE | PREGNANCY | LACTATION |
|---|---|---|---|---|
| 19 – 50 Years | 1.6g | 1.1g | 1.4g | 1.3g |
| 51+ Years | 1.6g | 1.1g | – | – |
In oncologic and inflammatory settings, higher therapeutic doses of omega-3 fatty acids – typically 2–4 g/day of combined EPA + DHA have been investigated as adjuncts to standard therapy for their potential to attenuate systemic inflammation, preserve lean body mass, and improve treatment tolerance. In a multicenter, double-blind randomized controlled trial that provided an EPA-enriched oral supplement delivering approximately 2.2 g EPA/day to patients with weight-losing gastrointestinal or lung cancer, improved weight maintenance and quality of life compared with isocaloric control was observed.104 In another RCT, patients receiving first-line chemotherapy for non-small-cell lung cancer, supplementation with fish oil providing approximately 2.2 g EPA + DHA per day improved chemotherapy completion rates and helped preserve lean body mass, compared with controls not receiving omega-3 fatty acids.105 In patients with metastatic colorectal cancer, the addition of fish oil to chemotherapy regimens was associated with a longer time to tumor progression and an improved overall treatment response rate, suggesting a potential synergistic effect between omega-3 fatty acids and cytotoxic therapy.106 Another study on colorectal cancer patients undergoing adjuvant chemotherapy, fish-oil supplementation led to a reduction in inflammatory markers (e.g., C-Reactive Protein) and an improvement in nutritional and performance status compared with non-supplemented controls.107
Reflecting the collective clinical evidence, the European Society for Clinical Nutrition and Metabolism (ESPEN) recommends that omega-3 supplementation be considered as part of a comprehensive, multimodal nutritional strategy for patients with cancer-related cachexia.108 Specifically, ESPEN suggests incorporating EPA at doses of approximately 1.5–2 g/day, either as a concentrated fish-oil supplement or as part of a specialized oral nutritional formula, to help stabilize appetite, body weight, and lean muscle mass when combined with adequate energy and protein intake. The ESPEN Practical Guideline (2021) further emphasizes that omega-3 supplementation should not be used in isolation, but rather as an adjunct to individualized nutrition counseling, physical activity, and optimal anti-cancer therapy.109
Table 6: RECOMMENDATIONS BASED ON RESEARCH
| USE/POPULATION | RECOMMENDED INTAKE | DON’T TAKE MORE THAN… | WHEN WILL I SEE RESULTS? |
|---|---|---|---|
| General adult population | ≥ 250 mg/day EPA + DHA | 3 g/day without supervision | 6 – 12 weeks (lipid or inflammatory markers) |
| Anti-inflammatory benefit | ~ 1 g/day | 3 g/day (medical supervision above this) | 4 – 8 weeks for inflammatory or lipid response |
| Cardiac benefit (CHD patients) | 1 g/day | 3 g/day | 8 – 12 weeks for lipid profile improvement |
| Cancer cachexia | ≥ 1.5 – 2 g/day EPA | 3 g/day unless supervised | 4 – 12 weeks for appetite and lean-mass stabilization |
| Cancer adjuvant therapy | 2 – 4 g/day | 5 g/day (upper safe limit) | 6 – 8 weeks for chemotherapy-tolerance benefits |
Facts and Misconceptions
Myth: “Eating a little fish or taking flaxseed oil provides enough omega-3s.”
Fact: While fatty fish and plant sources contribute beneficial fatty acids, they often do not provide sufficient EPA and DHA for optimal health. Plant oils such as flaxseed, chia, and walnuts provide α-linolenic acid (ALA), but the human conversion to long-chain forms is very limited—approximately 5–10% to EPA and 2–5% to DHA.110 Achieving cardioprotective or anti-inflammatory levels typically requires regular fatty-fish consumption (≥ 2 servings per week) or supplementation with pre-formed EPA and DHA.Myth: “Omega-3 supplements prevent all cancers.”
Fact: Evidence supports omega-3s as adjunctive or preventive, not curative. Large cohort studies such as the NIH–AARP Diet and Health Study and the Singapore Chinese Health Study suggest that higher omega-3 intake may reduce the risk of certain cancers, including breast and colorectal cancer. However, large RCTs such as VITAL and DO-HEALTH found no significant reduction in total cancer incidence with 1 g/day of EPA + DHA, though both confirmed long-term safety. Omega-3s may help modulate inflammation, support membrane function, and enhance treatment tolerance, but they do not replace standard oncologic therapy.Myth: “Omega-3s dangerously thin the blood.”
Fact: At typical dosages, omega-3s are safe and well tolerated. While very high doses can modestly prolong bleeding time, clinical bleeding events are rare. Supplemental intakes of EPA and DHA combined at doses up to about 5 g/day appear to be safe. These doses have not been shown to cause bleeding problems or affect immune function.111Myth: “More omega-3 is always better”
Fact: Higher doses do not necessarily produce stronger benefits and may increase the risk of mild side effects (e.g., nausea, reflux, or fishy aftertaste). Clinical advantages plateau around 1 g/day for cardiovascular protection and 2–4 g/day for therapeutic effects (e.g., cachexia, hypertriglyceridemia). Extremely high intakes (>4 g/day) have been linked to a slightly higher risk of atrial fibrillation in cardiovascular patients in large RCTs such as REDUCE-IT.112 Dose selection should match therapeutic goals.Myth: “Omega-3s work instantly.”
Fact: Omega-3s integrate gradually into cell membranes, and measurable benefits generally appear after 4 – 12 weeks of consistent intake.113 Improvements in triglyceride levels, inflammatory markers, or muscle mass typically require several months of sustained supplementation.
Food Sources
According to the National Institutes of Health (NIH) Office of Dietary Supplements, the richest dietary sources of omega-3 fatty acids are fatty fish and certain plant oils.114 The long-chain omega-3s eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) are found predominantly in marine sources, including salmon, mackerel, tuna, herring, sardines, and anchovies. However, certain large predatory fish such as king mackerel, shark, swordfish, and tilefish can contain elevated levels of methylmercury, which may pose health risks, particularly for pregnant and breastfeeding women, nursing infants, and young children. Evidence from seafood monitoring data shows that these high-trophic species consistently contain the highest methylmercury concentrations in U.S. markets.115 Smaller amounts of Omega-3 Fatty acids are present in other seafood, such as trout and oysters. The plant-based omega-3 α-linolenic acid (ALA) occurs mainly in flaxseed and flaxseed oil, chia seeds, canola oil, soybeans, soybean oil, walnuts, and hempseed. Below are two tables listing common food sources of omega-3 fatty acids.116
Table 7: PLANT-BASED SOURCES OF OMEGA-3 FATTY ACIDS
| FOOD | ALA (g) | DHA (g) | EPA (g) | |||||
|---|---|---|---|---|---|---|---|---|
| Flaxseed oil, 1 tbsp | 7.26 | — | — | |||||
| Chia seeds, 1 ounce | 5.06 | — | — | |||||
| English walnuts, 1 ounce | 2.57 | — | — | |||||
| Flaxseed, whole, 1 tbsp | 2.35 | — | — | |||||
| Canola oil, 1 tbsp | 1.28 | — | — | |||||
| Soybean oil, 1 tbsp | 0.92 | — | — | |||||
| Black walnuts, 1 ounce | 0.76 | — | — | |||||
| Mayonnaise, 1 tbsp | 0.74 | — | — | |||||
| Edamame, frozen, prepared, ½ cup | 0.28 | — | — | |||||
| Refried beans, canned, vegetarian, ½ cup | 0.21 | — | — | |||||
| Kidney beans, canned, ½ cup | 0.10 | — | — | |||||
| Baked beans, canned, vegetarian, ½ cup | 0.07 | — | — | |||||
| Bread, whole wheat, 1 slice | 0.04 | — | — | |||||
Table 8: ANIMAL-BASED SOURCES OF OMEGA-3 FATTY ACIDS
| FOOD | ALA (g) | DHA (g) | EPA (g) |
|---|---|---|---|
| Salmon, Atlantic, farmed, cooked, 3 oz | — | 1.24 | 0.59 |
| Salmon, Atlantic, wild, cooked, 3 oz | — | 1.22 | 0.35 |
| Herring, Atlantic, cooked, 3 oz | — | 0.94 | 0.77 |
| Sardines, canned in tomato sauce, 3 oz | — | 0.74 | 0.45 |
| Mackerel, Atlantic, cooked, 3 oz | — | 0.59 | 0.43 |
| Salmon, pink, canned, 3 oz | 0.04 | 0.63 | 0.28 |
| Trout, rainbow, wild, cooked, 3 oz | — | 0.44 | 0.40 |
| Oysters, eastern, wild, cooked, 3 oz | 0.14 | 0.23 | 0.30 |
| Sea bass, cooked, 3 oz | — | 0.47 | 0.18 |
| Shrimp, cooked, 3 oz | — | 0.12 | 0.12 |
| Lobster, cooked, 3 oz | 0.04 | 0.07 | 0.10 |
| Tuna, light, canned in water, 3 oz | — | 0.17 | 0.02 |
| Tilapia, cooked, 3 oz | 0.04 | 0.11 | — |
| Scallops, cooked, 3 oz | — | 0.09 | 0.06 |
| Cod, Pacific, cooked, 3 oz | — | 0.10 | 0.04 |
| Tuna, yellowfin, cooked, 3 oz | — | 0.09 | 0.01 |
| Ground beef, 85% lean, cooked, 3 oz | 0.04 | — | — |
| Egg, cooked, 1 | — | 0.03 | — |
| Chicken breast, roasted, 3 oz | — | 0.02 | 0.01 |
| Milk, low-fat (1%), 1 cup | — | 0.01 | — |
Fortified And Enriched Omega-3 Foods
Beyond natural marine and plant sources, several fortified foods now contribute meaningfully to omega-3 intake. The FAO/WHO Expert Consultation on Fats and Fatty Acids highlights fortification of dairy products, eggs, and spreads as a practical strategy to raise long-chain n-3 levels in populations with low fish consumption.117 Such fortification efforts aim to provide consistent sources of EPA and DHA in everyday foods, particularly in regions where fish consumption is limited or where dietary preferences favor vegetarian or plant-based patterns.
Several studies have demonstrated that enriching hen feed with fish oils or microalgae is an effective way to produce omega-3-enriched eggs containing significantly higher EPA and DHA levels. For example, eggs from hens fed omega-3-fortified diets were shown to contain up to 327 mg total n-3 PUFA per 100 g egg, including elevated EPA and DHA contents, relative to conventional eggs which contained 204 mg.118 These enriched eggs serve as one of the most accessible dietary sources of long-chain omega-3s, as fortification through feed supplementation directly enhances yolk lipid composition without compromising egg flavor or quality. Similarly, dairy milk can be successfully fortified with omega-3 fatty acids using microencapsulated fish oil without altering flavor or stability. In a controlled feeding trial, feeding cows linseed and fish oil has been shown to more than double milk’s omega-3 content.119 This demonstrates that both direct fortification (through addition of encapsulated oils) and indirect enrichment (through animal diet modification) are viable approaches for increasing omega-3 levels in dairy systems. Such methods not only enhance nutritional value but also maintain consumer acceptance of fortified milk. Fortifying dahi (Indian Yogurt) with microencapsulated flaxseed-oil powder increased α-linolenic acid (ALA) content total fatty acids, while preserving taste and stability during storage.120 Functional fermented dairy products like dahi and yogurt offer a convenient and culturally familiar medium for omega-3 incorporation, making them particularly suitable for widespread nutritional interventions in South Asian diets. Similarly, soy-, oat-, and almond-based drinks fortified with microalgal oil typically contain 40–80 mg DHA per 240 mL serving, offering a practical non-dairy option for increasing long-chain omega-3 intake.121 Because these plant-based beverages are often consumed daily, their fortification with microalgal oil provides an important vegan-friendly route to achieve recommended DHA intakes, bridging nutritional gaps for individuals who avoid fish or animal products.
Table 9: FORTIFIED AND ENRICHED OMEGA-3 FOODS
| FOOD/PRODUCT | FORTIFICATION/ENRICHMENT METHOD | MAIN OMEGA-3 TYPE(S) | OMEGA-3 CONTENT (PER SERVING OR 100 G) | NOTES |
|---|---|---|---|---|
| Omega-3 eggs | Hen feed enriched with fish oil or microalgae | EPA, DHA | Up to 327 mg total n-3 PUFA per 100 g (vs. 204 mg in regular eggs) | Fortification enhances yolk lipid composition without affecting flavor or texture |
| Fortified cow’s milk | Microencapsulated fish oil added directly; or cows fed linseed + fish oil | ALA, EPA, DHA | Omega-3 content more than doubled vs. regular milk | Maintains flavor and stability; suitable for large-scale dairy fortification |
| Fortified dahi (Indian yogurt) | Microencapsulated flaxseed-oil powder added | ALA | Significant rise in ALA % of total fatty acids | Taste and texture preserved; remains stable during cold storage |
| Fortified yogurt (general fermented dairy) | Microalgal or flaxseed oil incorporation | ALA, DHA | Variable; formulation-dependent | Functional fermented dairy foods are widely accepted vehicles for omega-3 delivery |
| Soy, oat, or almond beverages | Fortified with microalgal oil | DHA | 40–80 mg DHA per 240 mL serving | Provides a vegan-friendly, daily source of long-chain omega-3s |
| Fortified dairy systems (general) | Direct addition of encapsulated oils or indirect enrichment via animal diet | ALA, EPA, DHA | Variable, depending on feed and formulation | Enhances nutritional value while maintaining consumer acceptability |
Supplemental Sources
Omega-3 fatty acids are widely marketed in various supplement forms, including fish oil, krill oil, cod liver oil, and algal oil capsules designed for vegetarians.122 Fish-oil products typically contain around 1,000 mg of oil per capsule, though the proportion of active ingredients EPA and DHA varies considerably between brands. Most standard formulations provide roughly 180 mg of EPA and 120 mg of DHA per 1,000 mg capsule, while concentrated ‘high-strength’ fish-oil products may provide up to 500–600 mg combined EPA and DHA per soft gel. Cod liver oil combines omega-3s with naturally occurring vitamins A and D, making it a multifunctional option for consumers. Each 1,000 mg capsule of cod liver oil typically contains around 80–120 mg of EPA and a similar amount of DHA, though concentrations vary among brands. Despite concerns about mercury in seafood, purified omega-3 supplements are considered free of this contaminant because modern distillation and filtration processes effectively remove heavy metals.123 The chemical composition of omega-3 supplements differs by manufacturing method. They may be supplied as natural triglycerides, ethyl esters, re-esterified triglycerides, free fatty acids, or phospholipids. Triglyceride and re-esterified triglyceride forms resemble the structure found in whole fish and tend to be more efficiently absorbed than ethyl ester preparations, although all forms significantly raise circulating EPA and DHA. Controlled pharmacokinetic studies confirm that the molecular form influences absorption efficiency, with triglyceride and phospholipid formulations showing slightly higher bioavailability than ethyl esters124. Krill oil provides omega-3s largely in phospholipid form, which may improve absorption and promote more efficient incorporation of EPA and DHA into cell membranes.125 Common krill oil supplements provide 100–400 mg EPA and 50–200 mg DHA per capsule.126 On the other hand, Algal oil capsule supplements and algal oil-fortified foods are bioequivalent and safe sources of DHA, offering a clean, sustainable, and plant-based alternative for individuals who avoid fish or other marine sources.127 Plant-based sources of omega-3s from algal oil usually provide around 100–300 mg DHA. Because supplement formulations vary in concentration, purity, and chemical structure, consumers should review labels carefully to determine the exact amounts of EPA and DHA provided.
Table 10: OMEGA-3 FATTY ACID SUPPLEMENTS
| SUPPLEMENT TYPE | MAIN OMEGA-3 FORM(S) | TYPICAL EPA (MG) | TYPICAL DHA (MG) | OTHER COMPONENTS / NOTES |
|---|---|---|---|---|
| Fish oil (standard) | Triglycerides, ethyl esters, or re-esterified triglycerides | ~180 mg | ~120 mg | ~1,000 mg oil per capsule; active content varies by brand |
| Fish oil (high strength / concentrated) | Re-esterified triglycerides or ethyl esters | 250–400 mg | 250–300 mg | Up to 500–600 mg total EPA + DHA per softgel |
| Cod liver oil | Triglycerides | 80–120 mg | 80–120 mg | Also provides vitamins A & D; concentrations vary by brand |
| Krill oil | Phospholipids | 100–400 mg | 50–200 mg | High bioavailability due to phospholipid form; supports membrane incorporation |
| Algal oil (vegan) | Triglycerides or phospholipids | – | 100–300 mg | Plant-based, mercury-free DHA source; sustainable marine alternative |
| General purified omega-3 supplements | Triglycerides, ethyl esters, re-esterified triglycerides, free fatty acids, phospholipids | Variable | Variable | Purification removes mercury and heavy metals; bioavailability differs by form |
Side Effects
Omega-3 fatty acid supplements are generally recognized as safe and well-tolerated, though some individuals experience mild gastrointestinal discomfort such as nausea, diarrhea, or a fishy aftertaste. These effects are dose-dependent and usually improved by taking capsules with meals or using enteric-coated formulations.128 Other commonly reported adverse events include dyspepsia, bloating, eructation (fishy burps), fatigue, nasopharyngitis, and arthralgia, as noted in large post-marketing studies and reviews.129 Because these reactions are typically transient and dose-related, adjusting intake or formulation often restores tolerance without discontinuation.
At higher doses, particularly above 3 g/day of combined EPA and DHA, omega-3s can inhibit platelet aggregation and slightly prolong bleeding time; however, meta-analyses indicate that this effect rarely translates into clinically significant bleeding events. A large pooled analysis of cardiovascular trials found that omega-3 polyunsaturated fatty acid (PUFA) supplementation did not increase overall bleeding risk, though high-dose purified EPA may contribute to a modest rise in bleeding tendency.130 Nonetheless, caution is advised in patients receiving concurrent anticoagulants or antiplatelet therapy, where monitoring for minor bruising or nosebleeds is prudent. Thus, while mild hemostatic effects can occur at pharmacologic doses, these are not generally a barrier to routine use in clinical practice. Another consistent finding in recent outcome trials is a small but measurable increase in atrial fibrillation risk, particularly among patients with existing cardiovascular disease who consumed doses above 1 g/day; the risk was about 1.5-fold higher at these pharmacologic levels compared to lower doses.131 This pro-arrhythmic signal appears to be dose-dependent rather than compound-specific, highlighting the importance of individualized dosing and rhythm monitoring in high-risk cardiac patients. Lipid-related effects are also dose- and formulation-dependent: DHA-containing preparations can modestly raise LDL-cholesterol (often with a shift to larger, less atherogenic particles), whereas EPA-only products generally do not.132 Importantly, the triglyceride-lowering benefits of omega-3s typically outweigh these mild lipid alterations, especially when therapy is targeted toward cardiovascular risk reduction. In rare cases, high intakes have been linked to slight alterations in immune cell signaling and reduced pathogen defense in experimental models, suggesting that excessive long-term use may influence immune responses.133 Additional mild adverse reactions occasionally reported include transient headache, skin irritation, or changes in body odor due to oxidation of polyunsaturated fats. These effects remain uncommon and are generally reversible upon dose reduction.
Cod-liver oil formulations, though nutritionally rich, also contain vitamins A and D, and chronic high intake can cause hypervitaminosis A, particularly in pregnancy.134 As with all supplements, sourcing from reputable manufacturers ensures quality, purity, and accurate labeling of EPA and DHA concentrations. Overall, omega-3 supplementation remains one of the most extensively studied and safest nutritional interventions when used within recommended limits. Clinicians should tailor dosing to therapeutic goals and monitor for gastrointestinal intolerance, lipid shifts, bleeding tendencies, and arrhythmic symptoms at higher pharmacologic doses.
Table 11: ADVERSE EFFECTS AND SAFETY PROFILE OF OMEGA-3 FATTY ACID SUPPLEMENTATION
| CATEGORY | ADVERSE EFFECT / OBSERVATION | MECHANISM OR RISK FACTOR | CLINICAL SIGNIFICANCE / MANAGEMENT |
|---|---|---|---|
| Gastrointestinal | Nausea, diarrhea, dyspepsia, fishy aftertaste, bloating, halitosis | Fat digestion & oxidation products | Take with meals; use enteric-coated capsules |
| General | Fatigue, headache, nasopharyngitis, arthralgia | Mild systemic inflammatory response | Typically self-limited |
| Hematologic | Mild platelet inhibition, prolonged bleeding time | EPA/DHA interfere with thromboxane A₂ | Monitor if on anticoagulants; rarely clinically significant |
| Cardiac | Slight ↑ risk of atrial fibrillation at >1 g/day | Dose-dependent membrane effects on atrial cells | Monitor rhythm in CVD patients; risk ≈1.5× |
| Lipid Profile | Mild ↑ LDL-C with DHA formulations | Altered VLDL metabolism | Offset by ↓ triglycerides and ↑ particle size |
| Immune | Reduced host defense (experimental) | Excessive EPA/DHA dampen immune signaling | Avoid megadoses (>5 g/day) long term |
| Dermatologic / Misc. | Skin irritation, odor change | Lipid oxidation byproducts | Rare; reversible |
| Allergic | Reactions in fish/shellfish-sensitive individuals | Protein contamination | Use algal DHA instead |
| Vitamin Toxicity (Cod Liver Oil) | Hypervitaminosis A (esp. pregnancy) | Excess vitamin A accumulation | Use moderate doses or switch to purified fish oil |
| Contaminants | Mercury, PCBs, dioxins (removed by purification) | Environmental | Negligible in purified supplements |
Legal Status and Availability
Omega-3 fatty acids are legally sold in the United States as dietary supplements, not as prescription drugs. The FDA recognizes certain omega-3 ingredients (e.g., fish oil concentrates and DHA-rich algal oils) as Generally Recognized as Safe (GRAS) for use in foods and supplements, so they’re widely available over the counter as softgels, liquids, gummies, and in fortified foods.135 However, these products are not FDA-approved to treat or prevent disease, and their EPA and DHA content and purity may vary because they are regulated as dietary supplements under federal law, not as pharmaceuticals.136 By contrast, several prescription formulations such as icosapent ethyl (Vascepa®) have undergone full FDA review and are approved for managing severe hypertriglyceridemia.137
Bottom Line
Omega-3 fatty acids are essential nutrients that support heart, brain, and immune health by maintaining anti-inflammatory balance and healthy lipid metabolism. They are generally safe and well tolerated, with mild digestive discomfort being the most common side effect. Risks such as bleeding or irregular heartbeat occur mainly with excessive intake. Regular consumption through fish, fortified foods, or algal oil supplements can help meet daily needs, but quality and purity vary, so choosing reputable, third-party-tested products is recommended.
Specific Nutrient: Sulforaphane
What is it?
The National Cancer Institute (NCI) defines sulforaphane as a compound isolated from cruciferous vegetables that exhibits chemopreventive, antioxidant, and anti-inflammatory activity by stimulating detoxification enzymes and promoting the removal of reactive oxygen species.138 Broccoli sprouts are the richest known dietary source, containing 50 to 100 times more glucoraphanin, the direct precursor of sulforaphane than mature broccoli.139 Sulforaphane belongs to the isothiocyanate family of phytochemicals, a group of naturally occurring compounds responsible for the distinctive sharp aroma and mildly bitter flavor found in cruciferous vegetables such as broccoli, kale, cabbage, cauliflower, Brussels sprouts, collard greens, mustard greens, arugula, and radishes.140 The compound’s name is derived from “sulfur,” reflecting both its chemical composition and the characteristic scent released when these vegetables are cooked.
Interest in sulforaphane intensified after it was first isolated and identified as a potent inducer of protective enzymes from broccoli by researchers at Johns Hopkins University in 1992.141 Since then, extensive experimental and clinical studies have investigated its biological effects, establishing sulforaphane as a multi-targeted dietary compound that supports detoxification, antioxidant defense, and anti-inflammatory activity. Beyond its chemopreventive properties, growing evidence suggests potential benefits for neuroprotection, cardiometabolic health, and metabolic regulation, highlighting sulforaphane as one of the most promising plant-derived nutrients in chronic disease prevention.142
How does it work?
Sulforaphane is not found directly in foods; instead, cruciferous vegetables contain its precursor glucoraphanin, a type of glucosinolate. When plant cells are damaged through chopping, chewing, or mild cooking the enzyme myrosinase converts glucoraphanin into sulforaphane.143
Figure 8: Sulforaphane generation from Glucoraphanin
(A) Cruciferous vegetables are a rich source of glucoraphanin.
(B) Upon chewing or chopping, the myrosinase enzyme present in plant tissues or intestinal flora catalyzes the breakdown of glucoraphanin to SFN (Sulforaphane)
(C) SFN consequently becomes available to exert health benefits.
Source: https://www.mdpi.com/1420-3049/28/19/6902
Sulforaphane works primarily by strengthening the body’s natural defense systems at the cellular level. Instead of acting directly as an antioxidant like vitamin C or E, it stimulates the body to produce its own protective enzymes that neutralize harmful molecules and enhance detoxification. This long-lasting defense mechanism helps cells better manage oxidative stress, the imbalance between free radicals and antioxidants that contributes to inflammation, DNA damage, and cancer development.
A primary mode of action involves activation of the Nrf2–ARE (nuclear factor erythroid 2–related factor 2–antioxidant response element) pathway, which serves as a master regulator of cellular antioxidant and detoxification responses. Under normal conditions, Nrf2 is held inactive in the cytoplasm by the protein Keap1, which promotes its degradation. When sulforaphane enters the cell, it reacts with specific cysteine residues on Keap1, breaking this interaction and allowing Nrf2 to move into the nucleus.144 Once there, Nrf2 forms a heterodimer with small Maf (sMaf) proteins, and this complex binds to antioxidant response elements (AREs) in DNA and triggers the expression of multiple phase II detoxification and antioxidant enzymes, which collectively strengthen the cell’s ability to eliminate toxins and resist oxidative injury.145 This includes upregulation of glutathione-related enzymes, which boost glutathione synthesis and enhance conjugation and removal of carcinogens. Together, these antioxidant and detoxification pathways act as the body’s first line of defense, preventing the accumulation of oxidative stress.
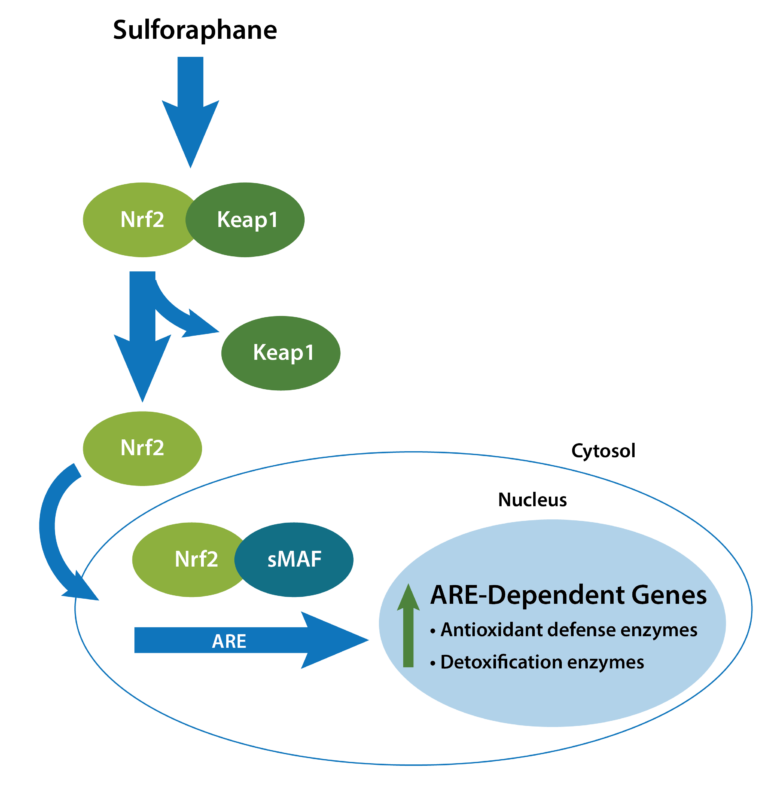
Figure 9: Activation of the Nrf2–Keap1–ARE pathway by Sulforaphane
Source: https://www.metagenicsinstitute.com/blogs/sulforaphane-nrf2-detoxification/
In addition to activating antioxidant defenses through Nrf2, sulforaphane also helps reduce inflammation by blocking the NF-κB (nuclear factor kappa-B) pathway.146 Normally, NF-κB controls the expression of inflammatory genes, but sulforaphane interferes with its activation, leading to lower production of inflammatory molecules such as IL-6, TNF-α, and COX-2. By modulating both Nrf2 and NF-κB, sulforaphane achieves a coordinated balance between antioxidant protection and inflammatory control, a hallmark of redox homeostasis (cellular balance of free radicals and antioxidants). This dual action, which is enhancing antioxidant enzymes while suppressing inflammatory signals, helps maintain cellular balance and protects tissues from chronic inflammation.
Sulforaphane further supports cellular health through epigenetic regulation. It acts as a natural histone deacetylase (HDAC) inhibitor, a mechanism that helps “unlock” silenced tumor-suppressor genes and restore normal gene activity.147 This modification of chromatin structure allows cells to maintain normal growth control and respond more effectively to stress signals. By modulating HDAC enzymes, sulforaphane can slow abnormal cell growth, encourage differentiation, and improve the cell’s ability to repair DNA damage. This epigenetic effect, along with its antioxidant and anti-inflammatory actions, underscores sulforaphane’s role as a broad-spectrum protector against chronic disease.
Beyond redox and epigenetic control, sulforaphane promotes apoptosis (programmed cell death) and cell-cycle arrest in cancer cells. It increases intracellular reactive oxygen species (ROS) generation, which activates pro-apoptotic pathways involving caspase-3 and caspase-9, while suppressing anti-apoptotic proteins such as Bcl-2 and survivin.148 This selective oxidative stress triggers mitochondrial dysfunction and halts the uncontrolled proliferation of tumor cells without harming normal cells, which possess stronger antioxidant defenses.
Figure 9: Epigenetic Mechanisms of Sulforaphane.149
Emerging research suggests that sulforaphane plays a significant role in maintaining gut homeostasis and gut–organ communication.150 It supports a balanced gut microbiota, enhances intestinal barrier integrity, and mitigates oxidative and inflammatory stress within the gut epithelium. By activating Nrf2 signaling and suppressing NF-κB–mediated inflammation, sulforaphane helps maintain mucosal immunity and promotes a favorable microbial environment. These actions contribute to systemic antioxidant balance and reduced risk of gut-derived organ dysfunction such as hepatic and cardiovascular injury. Collectively, these findings highlight sulforaphane’s emerging role in restoring gut–organ axis balance and preventing inflammation-related metabolic disturbances.
Figure 10: Anti-cancer activity of Sulforaphane
Source: https://www.frontiersin.org/journals/oncology/articles/10.3389/fonc.2023.1168321/full
What does the research tell us?
Human studies on sulforaphane have explored its roles in detoxifying carcinogens, reducing chronic inflammation, and modulating early cancer progression. Research has been conducted across Asia, the United States, and Europe, providing a broad clinical foundation relevant to inflammation-driven carcinogenesis.
One of the most important randomized clinical trials was carried out in 2014 in Qidong, China, a region with high exposure to airborne pollutants (benzene, acrolein, airborne aldehydes, combustion byproducts). In this 12-week randomized community trial, daily broccoli-sprout beverage consumption led to significant increases in urinary benzene (+61%) and acrolein (+23%) mercapturic acids demonstrating accelerated glutathione-mediated elimination of carcinogens.151 To date, this remains one of the strongest demonstrations of sulforaphane’s detoxification capacity in humans.
Sulforaphane has also been evaluated for its effects on infection-driven gastric inflammation, a key contributor to gastric cancer. In Japan, a controlled clinical trial in Helicobacter pylori infected adults in the year 2009, found that daily broccoli-sprout intake reduced gastric oxidative stress markers and mucosal inflammation, indicating disruption of a major chronic inflammation to cancer pathway.152 These early clinical trials helped establish sulforaphane’s relevance across different tissue types affected by inflammatory stress.
High-quality clinical evidence for sulforaphane’s chemopreventive potential in breast tissue comes from a 2015 human supplementation trial conducted in the United States.153 In women scheduled for breast biopsy, short-term supplementation with a standardized broccoli-sprout extract resulted in dose-dependent accumulation of sulforaphane metabolites in both plasma and breast tissue. The intervention increased the expression of key phase II cytoprotective enzymes, including NAD(P)H quinone oxidoreductase 1 (NQO1), which detoxifies reactive quinones and prevents free radical formation, and heme oxygenase-1 (HO-1), a stress-responsive enzyme that protects against oxidative injury and inflammation. The study also demonstrated significant in vivo histone deacetylase (HDAC) inhibition, supporting sulforaphane’s proposed epigenetic anticancer mechanism within human breast tissue. Together, these findings provided the first direct evidence that sulforaphane reaches human target tissues and induces molecular changes consistent with cancer prevention.
Sulforaphane has also been studied in men with recurrent or biochemically relapsing prostate cancer. The first was a phase II, single-arm clinical trial in men with recurrent prostate cancer who received sulforaphane-rich broccoli sprout extracts (200 µmol/day) for up to 20 weeks.154 The intervention was safe and well tolerated and found that sulforaphane supplementation slowed the rate of PSA rise and increased expression of Nrf2-dependent cytoprotective genes, although the primary endpoint of ≥50% PSA decline was rarely met. Building on this, a double-blind, randomized, placebo-controlled trial evaluated oral sulforaphane in men with biochemical recurrence after radical prostatectomy.155 Over 6 months of treatment, men in the sulforaphane group showed a significantly smaller rise in PSA and an 86% longer PSA doubling time compared with placebo (28.9 vs. 15.5 months), indicating slower biochemical progression. These clinical findings remain among the strongest human data supporting sulforaphane’s ability to modulate early cancer progression.
More recent work has focused on sulforaphane’s effects on the gut–organ axis, which is increasingly recognized as a driver of systemic inflammation and cancer risk.156 A 2023 hypothesis and mechanistic review outlined how sulforaphane may support gut homeostasis by activating Nrf2, reinforcing epithelial barrier integrity, reducing NF-κB–mediated inflammatory signaling, and modulating the gut microbiota in ways that lower oxidative and inflammatory stress along the gut–organ axis. These mechanisms are proposed to reduce downstream risk to organs such as the liver, cardiovascular system, and potentially cancer-prone tissues.
Several ongoing clinical trials are now exploring sulforaphane’s potential in cancer prevention and early-stage disease. Building on prior mechanistic and early human findings, these studies aim to determine whether sulforaphane’s molecular effects translate into clinically meaningful outcomes. A Phase II randomized clinical trial is evaluating whether sulforaphane can reduce bronchial epithelial proliferation in former smokers at high risk for lung cancer.157 The study measures changes in Ki-67 (a protein marker used to measure cell proliferation, meaning how fast cells are dividing) expression in bronchial biopsies as the primary endpoint, along with secondary biomarkers of oxidative stress and inflammation. In parallel, researchers are testing sulforaphane in other early-stage or high-risk settings. Another Phase II randomized clinical trial is investigating whether oral sulforaphane (Prostaphane®) can slow progression or recurrence in patients with non–muscle invasive bladder cancer.158 Participants receive sulforaphane or placebo following standard therapy, and the study tracks changes in recurrence intervals, tumor grade, and urinary biomarkers linked to oxidative stress, detoxification pathways, and inflammation. Extending this prevention-focused approach to dermatologic oncology, a Phase II prevention study is testing sulforaphane in individuals with multiple atypical nevi and a prior history of melanoma.159 The trial aims to determine whether daily sulforaphane supplementation can reduce the total area or number of atypical, pigmented lesions over 12 months, with secondary outcomes including dermoscopic changes and inflammatory biomarkers.
Table 12: ONGOING TRIALS ON SULFORAPHANE
| TRIAL FOCUS | POPULATION / SETTING | INTERVENTION | PRIMARY / SECONDARY ENDPOINTS |
|---|---|---|---|
| Lung Cancer Prevention (Phase II RCT) | Former smokers at high risk for lung cancer | Oral sulforaphane supplementation | Primary: Change in Ki-67 expression in bronchial epithelium. Secondary: Oxidative stress biomarkers, inflammatory markers |
| Bladder Cancer Recurrence Prevention (Phase II RCT) | Patients with non–muscle invasive bladder cancer after standard therapy | Oral Prostaphane® (sulforaphane) vs. placebo | Primary: Recurrence interval, tumor grade. Secondary: Urinary biomarkers (oxidative stress, detoxification, inflammation) |
| Melanoma Risk Reduction (Phase II Prevention Study) | Individuals with multiple atypical nevi and prior melanoma history | Daily sulforaphane supplementation | Primary: Change in total area/number of atypical, pigmented lesions. Secondary: Dermoscopic changes, inflammatory biomarkers |
In summary, human and preclinical studies consistently show that sulforaphane activates detoxification pathways, reduces inflammatory signaling, and influences early cancer biology across multiple tissues. While large long-term cancer-outcome trials are still needed, the strength and reproducibility of these biomarker and early-progression findings make sulforaphane one of the most promising dietary compounds under investigation for chronic inflammation and cancer prevention. Ongoing Phase II trials will help determine whether these molecular effects translate into measurable clinical benefit.
Dosage Recommendations Based on Research
Unlike omega-3 fatty acids or vitamins, sulforaphane does not have formal Dietary Reference Intakes (DRIs) or Adequate Intakes. Current dosing guidance comes almost entirely from human intervention trials using broccoli sprouts, sprout beverages, or standardized extracts, as well as pharmacokinetic and bioavailability studies. These studies consistently show that effective exposure depends not only on the nominal dose but also on the amount of glucoraphanin (the precursor), myrosinase activity, and individual differences in gut metabolism.160
Clinical detoxification studies show that doses providing ~50–150 μmol/day of sulforaphane equivalents are sufficient to activate glutathione-dependent detoxification pathways. In the Qidong randomized trial, participants consuming a broccoli-sprout beverage delivering ~150 μmol/day of combined sulforaphane/glucoraphanin experienced significant increases in urinary benzene and acrolein detoxification products, demonstrating robust activation of phase II detoxification.161
Comparable intakes delivered through sulforaphane-rich broccoli sprouts have been used in infection-driven gastric inflammation. In Helicobacter pylori–infected adults, daily consumption of 70 g of broccoli sprouts (containing several hundred micromoles of glucoraphanin) for 8 weeks reduced gastric oxidative stress markers and mucosal inflammation, supporting a detoxification/anti-inflammatory effect at physiologic, food-based doses.162
For tissue-level chemoprevention, lower but repeated doses have been effective. In women scheduled for breast biopsy, short-term supplementation with standardized broccoli-sprout extract providing tens of micromoles per day of sulforaphane equivalents led to detectable sulforaphane metabolites in plasma and breast tissue, induction of NQO1 and HO-1, and in vivo HDAC inhibition showing that relatively modest daily exposures can reach target tissue and trigger key anticancer pathways.163
Higher intakes have been tested in men with recurrent or biochemically relapsing prostate cancer. Phase II studies using sulforaphane-rich broccoli sprout extracts at the upper end of the nutritional range (on the order of 100–200 µmol/day of sulforaphane equivalents) over several months reported slowed PSA rise and prolongation of PSA-doubling time, without dose-limiting toxicity, suggesting that sustained higher nutritional doses may be appropriate in selected high-risk or early-disease settings.164
Systematic reviews also converge on similar effective ranges. A major review of clinical sulforaphane interventions noted that most human trials inducing measurable antioxidant and anti-inflammatory effects delivered between 25 and 200 μmol/day, with the 100–150 μmol/day range most consistently associated with improvements in oxidative stress markers, glutathione status, and inflammatory cytokines.165
A more recent review summarizing human sulforaphane trials across metabolic, pulmonary, and oncologic diseases reported that daily doses of ≈100–200 μmol reliably increased NQO1, HO-1, and other Nrf2 target genes while reducing IL-6, CRP, and TNF-α.166 The authors highlighted that this moderate range offers the optimal balance of efficacy, safety, and practicality for sustained use.
Together, these additional findings reinforce the conclusion that physiologic dosing in the 50–150 μmol/day range provides meaningful activation of detoxification and antioxidant pathways, whereas 100–200 μmol/day may be appropriate for targeted anti-inflammatory or chemopreventive purposes, particularly in controlled or clinical settings.
Table 13: RECOMMENDATIONS BASED ON RESEARCH
| USE/POPULATION | RECOMMENDED INTAKE | DON’T TAKE MORE THAN… | WHEN WILL I SEE RESULTS? |
|---|---|---|---|
| General Detoxification / Antioxidant support | 50–100 μmol/day | ~150 μmol/day (typical upper range used in detox trials) | 1–2 weeks (urinary detoxification markers) |
| Anti-Inflammatory benefit | 100–150 μmol/day | ~200 μmol/day (used in several clinical studies without toxicity) | 2–8 weeks (reductions in oxidative stress, IL-6, CRP; ↑ HO-1 / NQO1) |
| Targeted Chemoprevention | 100–200 μmol/day | ~200–300 μmol/day (short-term supervised use) | 4–12 weeks (Nrf2 gene induction; detox & inflammatory biomarkers improve) |
Facts and Misconceptions
Myth: “You need very high doses of sulforaphane for any real benefit.”
Fact: Sulforaphane is not a chemotherapeutic agent. Its primary role is modulating pathways involved in cancer risk including Nrf2 activation, HDAC inhibition, and NF-κB suppression. It supports prevention and early-stage modulation, not tumor eradication.Myth: “Raw broccoli gives you the same sulforaphane as sprouts or supplements.”
Fact: Raw broccoli contains the precursor glucoraphanin, but sulforaphane formation depends on myrosinase, which is largely destroyed by typical cooking methods. Broccoli sprouts contain 20–100× more glucoraphanin than mature broccoli, and consistently generate higher sulforaphane levels in humans.Myth: “Cooking destroys sulforaphane completely.”
Fact: Standard boiling does inactivate myrosinase, but sulforaphane can still form via gut microbiota conversion only at a reduced efficiency.167 Light steaming preserves up to 3–4× more sulforaphane potential compared with boiling.Myth: “Sulforaphane is only relevant for cancer.”
Fact: Sulforaphane’s strongest evidence spans detoxification, inflammation reduction, metabolic health, and gut–organ axis regulation, not just oncology.168 Reviews highlight its broad Nrf2-driven cytoprotective effects across multiple chronic diseases.
Food Sources
Among all cruciferous vegetables, broccoli sprouts are the richest natural source and deliver dramatically higher levels of glucoraphanin than mature broccoli or other Brassica vegetables. Sulforaphane can be generated from a wide variety of cruciferous vegetables, including broccoli, broccoli sprouts, Brussels sprouts, cabbage, cauliflower, kale, bok choy, collard greens, mustard greens, and watercress.169 Cruciferous vegetables remain the primary natural sources. Brassica species such as broccoli, broccoli sprouts, Brussels sprouts, kale, cabbage, bok choy, collard greens, and mustard greens contain variable glucoraphanin levels depending on cultivar, growing conditions, and preparation method. Broccoli sprouts, in particular, contain 20–100 times more glucoraphanin than mature broccoli heads.170 Other Brassica vegetables, including Brussels sprouts, cabbage, cauliflower, kale, bok choy, mustard greens, collard greens, and watercress also contain varying levels of glucosinolates. Cooking method plays a major role in sulforaphane availability. Myrosinase is heat-sensitive, and boiling or prolonged high-heat cooking can significantly reduce sulforaphane yield by inactivating the enzyme.171 Chemical studies show marked degradation of glucosinolates and diminished conversion when broccoli is boiled or microwaved excessively. In contrast, light steaming for 1–3 minutes preserves most myrosinase activity and maintains much higher sulforaphane-forming potential compared with boiling. Even when cooking destroys plant myrosinase, the human gut microbiota can convert glucoraphanin to sulforaphane, although with much lower and highly variable efficiency.172 This variability was demonstrated in controlled human studies showing wide individual differences in internal sulforaphane dose after ingesting the same amount of glucoraphanin. Importantly, sulforaphane formation from cooked broccoli can be restored 2–4× by adding raw myrosinase-containing foods such as mustard seeds, daikon radish, arugula, or sprouts.173
Table 14: SULFORAPHANE & GLUCOSINOLATE CONTENT OF COMMON CRUCIFEROUS VEGETABLES (PER 100 G RAW) 174
| VEGETABLE | GLUCOSINOLATES (MG) | SULFORAPHANE (µG) |
|---|---|---|
| Broccoli | 1297 mg | 260 µg |
| Cabbage | 1069 mg | 10.1 µg |
| Cauliflower | 1178 mg | ND |
| Brussels sprouts | 1013 mg | 2.6 µg |
| Kale | 1206 mg | 1736–3027 µg* |
| Arugula | 190 mg | 110 µg |
| Chinese cabbage | 297 mg | 540 µg |
| Collards | 2323 mg | ND |
| Horseradish | 11.4–36.4 mg | ND |
| Radishes | 676 mg | 16 µg |
| Rutabagas | 458 mg | 120 µg |
| Turnips | 698 mg | 60 µg |
| Watercress | 6–28 mg | ND |
| Kohlrabi | 829 mg | ND |
Supplemental Sources
Sulforaphane supplements vary widely in potency and bioavailability, largely because sulforaphane itself is unstable and difficult to deliver in its active form. Most commercial products provide glucoraphanin (GR), the precursor often without active myrosinase, resulting in highly variable and often low conversion to sulforaphane. Human pharmacokinetic studies show that conversion efficiency for GR-only preparations can be as low as 5–20% and varies more than 40-fold between individuals due to differences in gut microbiota.175 To overcome this variability, myrosinase-active supplements, typically broccoli-sprout powders or extracts retaining the enzyme, produce far higher and more consistent sulforaphane levels. Human feeding studies using sprouts or myrosinase-active beverages reported a several-fold increase in sulforaphane absorption and urinary dithiocarbamate recovery.176 These findings established that myrosinase presence is the single strongest determinant of sulforaphane bioavailability. Standardized, stabilized supplements have also been developed to provide more reliable dosing. One such formulation is Prostaphane®, a French pharmacy-grade sulforaphane preparation that demonstrated reproducible plasma sulforaphane levels and induction of Nrf2-target genes in clinical studies.177 In men with biochemical recurrence of prostate cancer, Prostaphane® significantly prolonged PSA-doubling time and reduced PSA rise over 6 months.178 To further improve stability and delivery, newer formulations use cyclodextrin inclusion complexes or liposomal encapsulation, which protect sulforaphane from degradation and enhance absorption. Cyclodextrin-complexed sulforaphane demonstrated superior stability and improved release kinetics in preclinical testing.179 Liposomal sulforaphane showed improved chemical stability and antioxidant activity in vitro compared with free sulforaphane suggesting potential advantages for future clinical applications.180 Overall, supplemental sources of sulforaphane differ dramatically in their ability to deliver meaningful internal exposure. Myrosinase-active preparations, stabilized sulforaphane formulations, and sprout-derived extracts consistently outperform glucoraphanin-only products.
Table 15: COMPARISON OF SULFORAPHANE SUPPLEMENTAL FORMS
| SUPPLEMENT TYPE | WHAT IT CONTAINS | BIOAVAILABILITY |
|---|---|---|
| Glucoraphanin-only (GR) supplements | GR precursor, no active myrosinase | Low and highly variable (5–20% conversion; ~40-fold inter-individual variation) |
| Myrosinase-active broccoli sprout powders/extracts | GR + active myrosinase | High and consistent conversion; several-fold higher sulforaphane absorption |
| Fresh broccoli sprouts | Naturally high GR + endogenous myrosinase | Excellent natural bioavailability; highest sulforaphane output among foods |
| Prostaphane® (stabilized sulforaphane) | Stabilized sulforaphane-rich extract | Reproducible plasma levels; clinically validated induction of Nrf2-target genes |
| Cyclodextrin-complexed sulforaphane | Sulforaphane embedded in cyclodextrin | Enhanced chemical stability and improved release profile |
| Liposomal sulforaphane | Sulforaphane in phospholipid micelles | Improved stability and absorption compared with free sulforaphane |
| Sulforaphane “precursor + probiotic” blends | GR + probiotic strains supporting conversion | Experimental; potentially higher conversion depending on microbiome |
Side Effects
Sulforaphane is generally regarded as safe in human studies, including trials using broccoli sprouts, sprout extracts, and stabilized sulforaphane preparations. Across clinical interventions, the most frequently reported side effects are mild, transient, and dose-related. Gastrointestinal symptoms are the most common adverse effects. Human trials of broccoli-sprout beverages and extracts have reported gastric discomfort, bloating, gas, diarrhea, or mild nausea, particularly when consuming high-glucosinolate foods or concentrated extracts.181 These effects are usually short-lived and improve with dose reduction. Some individuals experience increased bowel motility or a noticeable “broccoli odor” in urine or sweat due to isothiocyanate metabolism, a benign effect also noted in controlled human metabolic studies.182
Sulforaphane may also interact with certain medications by influencing drug-metabolizing enzymes. A laboratory study found that sulforaphane altered the activity of detoxification enzymes and transporters involved in processing drugs such as furosemide, verapamil, and ketoprofen, suggesting potential changes in drug effectiveness or clearance.183 These interactions have not been linked to serious clinical events but highlight the need for caution in individuals taking multiple medications. This is particularly relevant for drugs with narrow therapeutic windows or those heavily dependent on hepatic metabolism.
Additionally, because cruciferous vegetables contain goitrogenic glucosinolates, very high intakes may transiently inhibit iodine uptake in the thyroid, although clinical studies show this effect is minimal at typical dietary or supplemental sulforaphane doses and is unlikely to cause dysfunction in individuals with adequate iodine status.184 Individuals with untreated hypothyroidism or marginal iodine intake should still monitor their response when consuming large quantities of cruciferous vegetables or high-dose supplements.
Legal Status and Availability
In the United States, sulforaphane is sold legally as a dietary supplement, not as an FDA-approved drug. Products containing sulforaphane or its precursor glucoraphanin are regulated under the Dietary Supplement Health and Education Act (DSHEA), meaning manufacturers do not need FDA pre-approval but are responsible for safety and labeling. Because supplements are not required to prove potency or bioavailability, sulforaphane content varies widely across brands. Importantly, no sulforaphane product is FDA-approved for the prevention or treatment of cancer or any medical condition.
Dietary Strategy & Lifestyle Factors
Dietary patterns exert profound influence on chronic inflammation, oxidative stress, and metabolic regulation, all of which are key biological mechanisms that shape cancer risk across the lifespan. Among all studied dietary models, the Mediterranean diet has shown the most consistent anti-inflammatory and anticancer benefits. Characterized by high intake of fruits, vegetables, legumes, whole grains, nuts, seeds, fish, and extra-virgin olive oil, and low intake of red meat and ultra-processed foods, this diet reduces systemic inflammatory markers including CRP, IL-6, and TNF-α, largely due to its abundance of polyphenols, monounsaturated fats, and omega-3 fatty acids.185 These nutrient-dense foods collectively enhance antioxidant capacity, improve endothelial function, and support gut microbial balance, which further contributes to lowered inflammation and improved immune regulation. Evidence from the large PREDIMED randomized trial demonstrated that individuals assigned to a mediterranean diet, supplemented with extra-virgin olive oil or nuts had significantly lower inflammatory biomarkers and improved cardiometabolic profiles, which are factors directly linked to cancer development and progression.186 Importantly, the trial also highlighted that dietary quality—not merely caloric intake plays a decisive role in modulating disease pathways. Meta-analyses also show that higher adherence to mediterranean diet is associated with lower incidence and mortality of colorectal, breast, and aerodigestive cancers, highlighting its protective role in modulating inflammatory and oxidative pathways central to tumorigenesis.187

Figure 11: Mediterranean Diet Pyramid
Source: https://www.healthyfood.com/advice/how-to-get-the-health-benefits-of-a-mediterranean-diet/
Beyond the mediterranean diet, whole-food, plant-forward dietary patterns rich in cruciferous vegetables, berries, whole grains, nuts, and legumes support gut microbiome diversity and increase short-chain fatty acid production (such as butyrate), which strengthens epithelial integrity and suppresses NF-κB signaling, thereby lowering chronic inflammation.188 These foods also promote metabolic homeostasis, stabilize blood glucose, and reduce lipid peroxidation, all of which contribute to long-term protection against inflammation-driven diseases. In contrast, Western dietary patterns high in ultra-processed foods, added sugars, saturated fats, and advanced glycation end products (AGEs) promote oxidative stress, dysbiosis, and insulin resistance, all of which heighten inflammatory activity and increase cancer susceptibility. For individuals with suspected food sensitivities, inflammatory bowel disease, IBS, or autoimmune conditions, elimination diets which temporarily remove common triggers such as gluten, dairy, soy, eggs, or food additives have shown reductions in inflammatory markers and symptom burden. For example, the Autoimmune Protocol (AIP) diet demonstrated significant decreases in fecal calprotectin and clinical symptoms in IBD patients.189 This personalized approach underscores the importance of recognizing individual variability in dietary responses, microbiome composition, and immune activation. Taken together, the evidence supports dietary strategies centered on whole foods, balanced fatty acid intake, microbiome-supportive fiber, avoidance of ultra-processed foods, and personalized identification of triggers as foundational in reducing chronic inflammation and lowering long-term cancer risk.
Who are the experts?
1. Jed W. Fahey, ScD
Jed W. Fahey is a nutritional biochemist known for pioneering work on cruciferous vegetables and chemoprotection. He formerly directed the Cullman Chemoprotection Center at Johns Hopkins and has published extensively on sulforaphane, detoxification enzymes, and phytochemical bioactivity in humans.
Key Works:
2. Paul Talalay, MD
Dr. Paul Talalay (1923–2019) was a world-renowned cancer pharmacologist and a founding figure in the field of dietary chemoprotection. As a longtime professor at Johns Hopkins, he helped identify sulforaphane and revealed its ability to activate protective cellular pathways against carcinogens.
Key Works:
Positions and Views of Worldwide Governmental Medical and Health Organizations
Worldwide medical and health organizations consistently emphasize the role of diet and lifestyle in reducing chronic inflammation and lowering cancer risk. Major medical associations, including the American Medical Association (AMA) and Academy of Medical Sciences (UK), highlight that diets rich in whole grains, fruits, vegetables, and omega-3 fatty acids support lower systemic inflammation and improve long-term disease outcomes, whereas ultra-processed foods and high-sugar diets are associated with metabolic dysfunction and chronic inflammatory disorders. Healthcare quality organizations such as the Agency for Healthcare Research and Quality (AHRQ) and the Institute for Healthcare Improvement (IHI) reinforce that evidence-based nutrition interventions, particularly those promoting plant-forward dietary patterns can reduce morbidity and enhance population health. Similarly, the National Institute for Health and Care Excellence (NICE, UK) has issued clinical guidance noting that healthy dietary patterns high in fiber and low in red and processed meats reduce the risk of several cancers, and NICE routinely incorporates diet-related recommendations into cancer-prevention and chronic-disease guidelines. Global governmental institutions also provide disease-specific dietary guidance: the World Health Organization (WHO) warns that unhealthy diets are among the top modifiable risk factors for chronic inflammation and cancer; the CDC, NIH, Public Health Agency of Canada, Australia Department of Health, New Zealand Ministry of Health, Ireland Department of Health, and Sweden’s Ministry of Health and Social Affairs all similarly affirm that maintaining a nutrient-dense, minimally processed diet plays a critical role in reducing inflammatory burden, supporting metabolic health, and preventing chronic diseases, including cancer. These publicly available guidance documents collectively underscore that improving diet quality remains a central, evidence-supported strategy for reducing chronic inflammation and promoting long-term cancer prevention.
Nonprofits and Private Organizations
Several nonprofit and private organizations play a central role in advancing public understanding of chronic inflammation, cancer risk, and the influence of diet on these processes. The American Cancer Society (ACS) provides accessible, evidence-based recommendations emphasizing diets rich in vegetables, fruits, whole grains, and omega-3–containing foods to reduce inflammation and support cancer prevention. The World Cancer Research Fund (WCRF) and the American Institute for Cancer Research (AICR) similarly highlight strong evidence linking dietary patterns low in processed foods and high in plant-based nutrients to reduce cancer incidence; both organizations publish regular expert reports on diet, inflammation, and cancer mechanisms. Disease-specific nonprofits such as the Crohn’s & Colitis Foundation and the Arthritis Foundation offer resources on anti-inflammatory dietary strategies aimed at reducing flare-ups and lowering systemic inflammatory burden. Many of these organizations maintain active social-media platforms and online communities that disseminate educational materials, infographics, and evidence summaries to millions of followers, making high-quality dietary guidance accessible to the public. Together, these nonprofits shape national discourse on diet and inflammation and support individuals seeking credible, research-based nutritional strategies for chronic disease prevention.
Bibliography
Ağagündüz, Duygu, Teslime Özge Şahin, Birsen Yılmaz, Kübra Damla Ekenci, Şehriban Duyar Özer, and Raffaele Capasso. 2022. “Cruciferous Vegetables and Their Bioactive Metabolites: From Prevention to Novel Therapies of Colorectal Cancer.” Evidence-Based Complementary and Alternative Medicine 2022 (1): 1534083. https://doi.org/10.1155/2022/1534083.
Aggarwal, Bharat B., and Kuzhuvelil B. Harikumar. 2009. “Potential Therapeutic Effects of Curcumin, the Anti-Inflammatory Agent, Against Neurodegenerative, Cardiovascular, Pulmonary, Metabolic, Autoimmune and Neoplastic Diseases.” The International Journal of Biochemistry & Cell Biology, Directed Issue: Epigenetics and Disease, 41 (1): 40–59. https://doi.org/10.1016/j.biocel.2008.06.010.
Aggarwal, V., H. Tuli, Ayşegül Varol, Falak Thakral, M. Yerer, K. Sak, M. Varol, Aklank Jain, Md Asaduzzaman Khan, and G. Sethi. 2019. “Role of Reactive Oxygen Species in Cancer Progression: Molecular Mechanisms and Recent Advancements.” Biomolecules 9. https://doi.org/10.3390/biom9110735.
Aguilar-Cazares, Dolores, Rodolfo Chavez-Dominguez, Angeles Carlos-Reyes, César Lopez-Camarillo, Olga N. Hernandez de la Cruz, and Jose S. Lopez-Gonzalez. 2019. “Contribution of Angiogenesis to Inflammation and Cancer.” Frontiers in Oncology 9 (December). https://doi.org/10.3389/fonc.2019.01399.
Ak, Tuba, and İlhami Gülçin. 2008. “Antioxidant and Radical Scavenging Properties of Curcumin.” Chemico-Biological Interactions 174 (1): 27–37. https://doi.org/10.1016/j.cbi.2008.05.003.
Alexander, Walter. 2013. “Prostate Cancer Risk and Omega-3 Fatty Acid Intake from Fish Oil: A Closer Look at Media Messages Versus Research Findings.” P & T: A Peer-Reviewed Journal for Formulary Management 38 (9): 561–64.
Alumkal, Joshi J., Rachel Slottke, Jacob Schwartzman, Ganesh Cherala, Myrna Munar, Julie N. Graff, Tomasz M. Beer, et al. 2015a. “A Phase II Study of Sulforaphane-Rich Broccoli Sprout Extracts in Men with Recurrent Prostate Cancer.” Investigational New Drugs 33 (2): 480–89. https://doi.org/10.1007/s10637-014-0189-z.
———, et al. 2015b. “A Phase II Study of Sulforaphane-Rich Broccoli Sprout Extracts in Men with Recurrent Prostate Cancer.” Investigational New Drugs 33 (2): 480–89. https://doi.org/10.1007/s10637-014-0189-z.
———, et al. 2015c. “A Phase II Study of Sulforaphane-Rich Broccoli Sprout Extracts in Men with Recurrent Prostate Cancer.” Investigational New Drugs 33 (2): 480–89. https://doi.org/10.1007/s10637-014-0189-z.
Anderson, Breanne M, and David WL Ma. 2009. “Are All n-3 Polyunsaturated Fatty Acids Created Equal?” Lipids in Health and Disease 8 (August): 33. https://doi.org/10.1186/1476-511X-8-33.
Angelo, Laura S., and Razelle Kurzrock. 2007. “Vascular Endothelial Growth Factor and Its Relationship to Inflammatory Mediators.” Clinical Cancer Research 13 (10): 2825–30. https://doi.org/10.1158/1078-0432.CCR-06-2416.
Arem, Hannah, and Erikka Loftfield. 2017. “Cancer Epidemiology: A Survey of Modifiable Risk Factors for Prevention and Survivorship.” American Journal of Lifestyle Medicine 12 (3): 200–210. https://doi.org/10.1177/1559827617700600.
Arends, Jann, Patrick Bachmann, Vickie Baracos, Nicole Barthelemy, Hartmut Bertz, Federico Bozzetti, Ken Fearon, et al. 2017. “ESPEN Guidelines on Nutrition in Cancer Patients.” Clinical Nutrition 36 (1): 11–48. https://doi.org/10.1016/j.clnu.2016.07.015.
Aronson, William J., Tristan Grogan, Pei Liang, Patricia Jardack, Amana R. Liddell, Claudia Perez, David Elashoff, et al. 2025. “High Omega-3, Low Omega-6 Diet With Fish Oil for Men With Prostate Cancer on Active Surveillance: The CAPFISH-3 Randomized Clinical Trial.” Journal of Clinical Oncology: Official Journal of the American Society of Clinical Oncology 43 (7): 800–809. https://doi.org/10.1200/JCO.24.00608.
Arterburn, Linda M., Harry A. Oken, James P. Hoffman, Eileen Bailey-Hall, Gloria Chung, Dror Rom, Jacqueline Hamersley, and Deanna McCarthy. 2007. “Bioequivalence of Docosahexaenoic Acid from Different Algal Oils in Capsules and in a DHA-Fortified Food.” Lipids 42 (11): 1011. https://doi.org/10.1007/s11745-007-3098-5.
Asif Ali, Muhammad, Noohela Khan, Nabeeha Kaleem, Waqas Ahmad, Salem Hussain Alharethi, Bandar Alharbi, Hassan H. Alhassan, et al. 2023. “Anticancer Properties of Sulforaphane: Current Insights at the Molecular Level.” Frontiers in Oncology 13 (June). https://doi.org/10.3389/fonc.2023.1168321.
Atwell, Lauren L., Zhenzhen Zhang, Motomi Mori, Paige Farris, John T. Vetto, Arpana M. Naik, Karen Y. Oh, Philippe Thuillier, Emily Ho, and Jackilen Shannon. 2015. “Sulforaphane Bioavailability and Chemopreventive Activity in Women Scheduled for Breast Biopsy.” Cancer Prevention Research (Philadelphia, Pa.) 8 (12): 1184–91. https://doi.org/10.1158/1940-6207.CAPR-15-0119.
Badmanaban, R., Dhananjoy Saha, Dhrubo J. Sen, Arpita Biswas, Supradip Mandal, and Susmita Basak. 2021. “Turmeric: A Holistic Solution for Biochemical Malfunction.” Research Journal of Pharmacy and Technology 14 (10): 5540–50. https://doi.org/10.52711/0974-360X.2021.00966.
Baenas, Nieves, Javier Marhuenda, Cristina García-Viguera, Pilar Zafrilla, and Diego A. Moreno. 2019. “Influence of Cooking Methods on Glucosinolates and Isothiocyanates Content in Novel Cruciferous Foods.” Foods 8 (7): 257. https://doi.org/10.3390/foods8070257.
Bahramsoltani, Roodabeh, Roja Rahimi, and Mohammad Hosein Farzaei. 2017. “Pharmacokinetic Interactions of Curcuminoids with Conventional Drugs: A Review.” Journal of Ethnopharmacology 209 (September): 1–12. https://doi.org/10.1016/j.jep.2017.07.022.
Beam, Ashley, Elizabeth Clinger, and Lei Hao. 2021. “Effect of Diet and Dietary Components on the Composition of the Gut Microbiota.” Nutrients 13 (8): 2795. https://doi.org/10.3390/nu13082795.
Bentley, StephenJ., DavidJ. Pearson, and KeithJ. B. Rix. 1983. “FOOD HYPERSENSITIVITY IN IRRITABLE BOWEL SYNDROME.” The Lancet, Originally published as Volume 2, Issue 8345, 322 (8345): 295–97. https://doi.org/10.1016/S0140-6736(83)90285-4.
Bhatt, Deepak L., P. Gabriel Steg, Michael Miller, Eliot A. Brinton, Terry A. Jacobson, Steven B. Ketchum, Ralph T. Doyle, et al. 2019. “Cardiovascular Risk Reduction with Icosapent Ethyl for Hypertriglyceridemia.” New England Journal of Medicine 380 (1): 11–22. https://doi.org/10.1056/NEJMoa1812792.
Bimonte, Sabrina, Antonio Barbieri, Giuseppe Palma, Domenica Rea, Antonio Luciano, Massimiliano D’Aiuto, Claudio Arra, and Francesco Izzo. 2015. “Dissecting the Role of Curcumin in Tumour Growth and Angiogenesis in Mouse Model of Human Breast Cancer.” BioMed Research International 2015: 878134. https://doi.org/10.1155/2015/878134.
Borm, Paul J. A., Paul Fowler, and David Kirkland. 2018. “An Updated Review of the Genotoxicity of Respirable Crystalline Silica.” Particle and Fibre Toxicology 15 (1): 23. https://doi.org/10.1186/s12989-018-0259-z.
Bosire, Claire, Meir J. Stampfer, Amy F. Subar, Yikyung Park, Sharon I. Kirkpatrick, Stephanie E. Chiuve, Albert R. Hollenbeck, and Jill Reedy. 2013. “Index-Based Dietary Patterns and the Risk of Prostate Cancer in the NIH-AARP Diet and Health Study.” American Journal of Epidemiology 177 (6): 504–13. https://doi.org/10.1093/aje/kws261.
Bouranis, John A, Laura M Beaver, Carmen P Wong, Jaewoo Choi, Sean Hamer, Ed W Davis, Kevin S Brown, et al. 2024. “Sulforaphane and Sulforaphane-Nitrile Metabolism in Humans Following Broccoli Sprout Consumption: Inter-Individual Variation, Association with Gut Microbiome Composition, and Differential Bioactivity.” Molecular Nutrition & Food Research 68 (4): e2300286. https://doi.org/10.1002/mnfr.202300286.
Braicu, Cornelia, Mihail Buse, Constantin Busuioc, Rares Drula, Diana Gulei, Lajos Raduly, Alexandru Rusu, et al. 2019. “A Comprehensive Review on MAPK: A Promising Therapeutic Target in Cancer.” Cancers 11 (10): 1618. https://doi.org/10.3390/cancers11101618.
Brasky, Theodore M., Amy K. Darke, Xiaoling Song, Catherine M. Tangen, Phyllis J. Goodman, Ian M. Thompson, Frank L. Meyskens, et al. 2013. “Plasma Phospholipid Fatty Acids and Prostate Cancer Risk in the SELECT Trial.” Journal of the National Cancer Institute 105 (15): 1132–41. https://doi.org/10.1093/jnci/djt174.
Brasky, Theodore M., Johanna W. Lampe, John D. Potter, Ruth E. Patterson, and Emily White. 2010. “Specialty Supplements and Breast Cancer Risk in the VITamins And Lifestyle (VITAL) Cohort.” Cancer Epidemiology, Biomarkers & Prevention: A Publication of the American Association for Cancer Research, Cosponsored by the American Society of Preventive Oncology 19 (7): 1696–1708. https://doi.org/10.1158/1055-9965.EPI-10-0318.
Buhrmann, Constanze, Ali Mobasheri, Franziska Busch, Constance Aldinger, Ralf Stahlmann, Azadeh Montaseri, and Mehdi Shakibaei. 2011. “Curcumin Modulates Nuclear Factor κB (NF-κB)-Mediated Inflammation in Human Tenocytes in Vitro.” The Journal of Biological Chemistry 286 (32): 28556–66. https://doi.org/10.1074/jbc.M111.256180.
Caires, Rebeca, Francisco J. Sierra-Valdez, Jonathan R. M. Millet, Joshua D. Herwig, Esra Roan, Valeria Vásquez, and Julio F. Cordero-Morales. 2017. “Omega-3 Fatty Acids Modulate TRPV4 Function Through Plasma Membrane Remodeling.” Cell Reports 21 (1): 246–58. https://doi.org/10.1016/j.celrep.2017.09.029.
Calder, Philip C. 2017. “Omega-3 Fatty Acids and Inflammatory Processes: From Molecules to Man.” Biochemical Society Transactions 45 (5): 1105–15. https://doi.org/10.1042/BST20160474.
———. n.d. “Dietary Factors and Low-Grade Inflammation in Relation to Overweight and Obesity Revisted.” The British Journal of Nutrition 127 (10): 1455–57. Accessed December 8, 2025. https://doi.org/10.1017/S0007114522000782.
Camargo, Carolina de Quadros, Michel Carlos Mocellin, Juliana de Aguiar Pastore Silva, Maria Emilia de Souza Fabre, Everson Araújo Nunes, and Erasmo Benicio Santos de Moraes Trindade. 2016. “Fish Oil Supplementation During Chemotherapy Increases Posterior Time to Tumor Progression in Colorectal Cancer.” Nutrition and Cancer 68 (1): 70–76. https://doi.org/10.1080/01635581.2016.1115097.
“Cancer.” n.d. Accessed July 6, 2025. https://www.who.int/news-room/fact-sheets/detail/cancer.
“Cancer Today.” n.d. Accessed July 6, 2025. https://gco.iarc.who.int/today/.
Cao, Yaning, Yanan Yi, Chongxu Han, and Bingwei Shi. 2024. “NF-κB Signaling Pathway in Tumor Microenvironment.” Frontiers in Immunology 15 (October). https://doi.org/10.3389/fimmu.2024.1476030.
Carroll, Robert E., Richard V. Benya, Danielle Kim Turgeon, Shaiju Vareed, Malloree Neuman, Luz Rodriguez, Madhuri Kakarala, et al. 2011. “Phase IIA Clinical Trial of Curcumin for the Prevention of Colorectal Neoplasia.” Cancer Prevention Research (Philadelphia, Pa.) 4 (3): 354–64. https://doi.org/10.1158/1940-6207.CAPR-10-0098.
Chevalier, Laurie, Annick Vachon, and Mélanie Plourde. 2021. “Pharmacokinetics of Supplemental Omega-3 Fatty Acids Esterified in Monoglycerides, Ethyl Esters, or Triglycerides in Adults in a Randomized Crossover Trial.” The Journal of Nutrition 151 (5): 1111–18. https://doi.org/10.1093/jn/nxaa458.
Cipolla, Bernard G., Eric Mandron, Jean Marc Lefort, Yves Coadou, Emmanuel Della Negra, Luc Corbel, Ronan Le Scodan, Abdel Rahmene Azzouzi, and Nicolas Mottet. 2015a. “Effect of Sulforaphane in Men with Biochemical Recurrence After Radical Prostatectomy.” Cancer Prevention Research 8 (8): 712–19. https://doi.org/10.1158/1940-6207.CAPR-14-0459.
———. 2015b. “Effect of Sulforaphane in Men with Biochemical Recurrence After Radical Prostatectomy.” Cancer Prevention Research 8 (8): 712–19. https://doi.org/10.1158/1940-6207.CAPR-14-0459.
Clarke, John D., Roderick H. Dashwood, and Emily Ho. 2008. “Multi-Targeted Prevention of Cancer by Sulforaphane.” Cancer Letters, Natural Products Special Issue, 269 (2): 291–304. https://doi.org/10.1016/j.canlet.2008.04.018.
Clarke, John, Anna Hsu, Kenneth Riedl, Deborah Bella, Steven Schwartz, Jan Stevens, and Emily Ho. 2011. “Bioavailability and Inter-Conversion of Sulforaphane and Erucin in Human Subjects Consuming Broccoli Sprouts or Broccoli Supplement in a Cross-over Study Design.” Pharmacological Research : The Official Journal of the Italian Pharmacological Society 64 (July): 456–63. https://doi.org/10.1016/j.phrs.2011.07.005.
Contributor, WebMD Editorial. n.d. “Healthy Foods High in Omega-6.” WebMD. Accessed October 22, 2025. https://www.webmd.com/diet/foods-high-in-omega-6.
Cruz-Correa, Marcia, Daniel A. Shoskes, Patricia Sanchez, Rhongua Zhao, Linda M. Hylind, Steven D. Wexner, and Francis M. Giardiello. 2006. “Combination Treatment with Curcumin and Quercetin of Adenomas in Familial Adenomatous Polyposis.” Clinical Gastroenterology and Hepatology: The Official Clinical Practice Journal of the American Gastroenterological Association 4 (8): 1035–38. https://doi.org/10.1016/j.cgh.2006.03.020.
Cruz–Correa, Marcia, Daniel A. Shoskes, Patricia Sanchez, Rhongua Zhao, Linda M. Hylind, Steven D. Wexner, and Francis M. Giardiello. 2006. “Combination Treatment With Curcumin and Quercetin of Adenomas in Familial Adenomatous Polyposis.” Clinical Gastroenterology and Hepatology 4 (8): 1035–38. https://doi.org/10.1016/j.cgh.2006.03.020.
Cuomo, John, Giovanni Appendino, Adam S. Dern, Erik Schneider, Toni P. McKinnon, Mark J. Brown, Stefano Togni, and Brian M. Dixon. 2011. “Comparative Absorption of a Standardized Curcuminoid Mixture and Its Lecithin Formulation.” Journal of Natural Products 74 (4): 664–69. https://doi.org/10.1021/np1007262.
“Curcumin and Quercetin Modify Warfarin-Induced Regulation of Porcine CYP1A2 and CYP3A Expression and Activity in Vitro: Xenobiotica: Vol 52 , No 5 – Get Access.” n.d. Accessed August 12, 2025. https://www.tandfonline.com/doi/full/10.1080/00498254.2022.2089932.
Daak, Ahmed A., Abozer Y. Elderdery, Leana M. Elbashir, Katia Mariniello, Jeremy Mills, Garry Scarlett, Mustafa I. Elbashir, and Kebreab Ghebremeskel. 2015. “Omega 3 (n − 3) Fatty Acids down-Regulate Nuclear Factor-Kappa B (NF-κB) Gene and Blood Cell Adhesion Molecule Expression in Patients with Homozygous Sickle Cell Disease.” Blood Cells, Molecules, and Diseases 55 (1): 48–55. https://doi.org/10.1016/j.bcmd.2015.03.014.
“Definition of Sulforaphane – NCI Drug Dictionary – NCI.” 2011. {nciAppModulePage}. https://www.cancer.gov/publications/dictionaries/cancer-drug/def/sulforaphane.
deMontalk, Jen. 2022. “How to Get the Health Benefits of a Mediterranean Diet.” Healthy Food Guide. https://www.healthyfood.com/advice/how-to-get-the-health-benefits-of-a-mediterranean-diet/.
Dinkova-Kostova, Albena T., W. David Holtzclaw, Robert N. Cole, Ken Itoh, Nobunao Wakabayashi, Yasutake Katoh, Masayuki Yamamoto, and Paul Talalay. 2002. “Direct Evidence That Sulfhydryl Groups of Keap1 Are the Sensors Regulating Induction of Phase 2 Enzymes That Protect Against Carcinogens and Oxidants.” Proceedings of the National Academy of Sciences of the United States of America 99 (18): 11908–13. https://doi.org/10.1073/pnas.172398899.
Djuricic, Ivana, and Philip C. Calder. 2021. “Beneficial Outcomes of Omega-6 and Omega-3 Polyunsaturated Fatty Acids on Human Health: An Update for 2021.” Nutrients 13 (7): 2421. https://doi.org/10.3390/nu13072421.
Dmytriv, Tetiana R., Oleh Lushchak, and Volodymyr I. Lushchak. 2025. “Glucoraphanin Conversion into Sulforaphane and Related Compounds by Gut Microbiota.” Frontiers in Physiology 16 (February). https://doi.org/10.3389/fphys.2025.1497566.
“DO-HEALTH.” 2025. https://do-health.eu/.
Dominguez, Edward C., Art J. Heires, Jacqueline Pavlik, Tricia D. Larsen, Stephanie Guardado, Joseph H. Sisson, Michelle L. Baack, Debra J. Romberger, and Tara M. Nordgren. 2020. “A High Docosahexaenoic Acid Diet Alters the Lung Inflammatory Response to Acute Dust Exposure.” Nutrients 12 (8): 2334. https://doi.org/10.3390/nu12082334.
“Drug Approval Package.” n.d. Accessed October 23, 2025. https://www.accessdata.fda.gov/drugsatfda_docs/nda/2012/202057Orig1s000TOC.cfm.
EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). 2012. “Scientific Opinion on the Tolerable Upper Intake Level of Eicosapentaenoic Acid (EPA), Docosahexaenoic Acid (DHA) and Docosapentaenoic Acid (DPA).” EFSA Journal 10 (7): 2815. https://doi.org/10.2903/j.efsa.2012.2815.
Egner, Patricia A., Jian-Guo Chen, Adam T. Zarth, Derek K. Ng, Jin-Bing Wang, Kevin H. Kensler, Lisa P. Jacobson, et al. 2014. “Rapid and Sustainable Detoxication of Airborne Pollutants by Broccoli Sprout Beverage: Results of a Randomized Clinical Trial in China.” Cancer Prevention Research 7 (8): 813–23. https://doi.org/10.1158/1940-6207.CAPR-14-0103.
ElKhalifa, Dana, Nour Al-Ziftawi, Ahmed Awaisu, Feras Alali, and Ashraf Khalil. 2023. “Efficacy and Tolerability of Sulforaphane in the Therapeutic Management of Cancers: A Systematic Review of Randomized Controlled Trials.” Frontiers in Oncology 13 (November). https://doi.org/10.3389/fonc.2023.1251895.
“Essential Fatty Acids Linus Pauling Institute Oregon State University.” 2014. https://lpi.oregonstate.edu/mic/other-nutrients/essential-fatty-acids.
Estruch, Ramón, Emilio Ros, Jordi Salas-Salvadó, Maria-Isabel Covas, Dolores Corella, Fernando Arós, Enrique Gómez-Gracia, et al. 2018. “Retraction and Republication: Primary Prevention of Cardiovascular Disease with a Mediterranean Diet. N Engl J Med 2013;368:1279-90.” New England Journal of Medicine 378 (25): 2441–42. https://doi.org/10.1056/NEJMc1806491.
“Excess Omega-3 Fatty Acids Could Lead to Negative Health Effects Newsroom Oregon State University.” 2013. https://news.oregonstate.edu/news/excess-omega-3-fatty-acids-could-lead-negative-health-effects.
Fabian, Carol J., Bruce F. Kimler, and Stephen D. Hursting. 2015. “Omega-3 Fatty Acids for Breast Cancer Prevention and Survivorship.” Breast Cancer Research 17 (1): 62. https://doi.org/10.1186/s13058-015-0571-6.
Fahey, J. W., Y. Zhang, and P. Talalay. 1997. “Broccoli Sprouts: An Exceptionally Rich Source of Inducers of Enzymes That Protect Against Chemical Carcinogens.” Proceedings of the National Academy of Sciences of the United States of America 94 (19): 10367–72. https://doi.org/10.1073/pnas.94.19.10367.
Fahey, Jed W., Amy T. Zalcmann, and Paul Talalay. 2001. “The Chemical Diversity and Distribution of Glucosinolates and Isothiocyanates Among Plants.” Phytochemistry 56 (1): 5–51. https://doi.org/10.1016/S0031-9422(00)00316-2.
Fahey, Jed W., Yuesheng Zhang, and Paul Talalay. 1997. “Broccoli Sprouts: An Exceptionally Rich Source of Inducers of Enzymes That Protect Against Chemical Carcinogens.” Proceedings of the National Academy of Sciences 94 (19): 10367–72. https://doi.org/10.1073/pnas.94.19.10367.
Fearon, K C H, M F von Meyenfeldt, A G W Moses, R van Geenen, A Roy, D J Gouma, A Giacosa, et al. 2003. “Effect of a Protein and Energy Dense n-3 Fatty Acid Enriched Oral Supplement on Loss of Weight and Lean Tissue in Cancer Cachexia: A Randomised Double Blind Trial.” Gut 52 (10): 1479–86. https://doi.org/10.1136/gut.52.10.1479.
“Figure 4. NF-κB Inhibition by Curcumin. Figure 4: Curcumin Prevents The…” n.d. ResearchGate. Accessed September 30, 2025. https://www.researchgate.net/figure/NF-kB-inhibition-by-curcumin-Figure-4-Curcumin-prevents-the-entry-of-NF-kB-into-nucleus_fig4_324650356.
Fioranelli, Massimo, Anna G. Bottaccioli, Francesco Bottaccioli, Maria Bianchi, Miriam Rovesti, and Maria G. Roccia. 2018. “Stress and Inflammation in Coronary Artery Disease: A Review Psychoneuroendocrineimmunology-Based.” Frontiers in Immunology 9 (September). https://doi.org/10.3389/fimmu.2018.02031.
“FISH OIL: Overview, Uses, Side Effects, Precautions, Interactions, Dosing and Reviews.” n.d. Accessed October 20, 2025. https://www.webmd.com/vitamins/ai/ingredientmono-993/fish-oil.
Gago-Dominguez, M., J.-M. Yuan, C.-L. Sun, H.-P. Lee, and M. C. Yu. 2003. “Opposing Effects of Dietary n-3 and n-6 Fatty Acids on Mammary Carcinogenesis: The Singapore Chinese Health Study.” British Journal of Cancer 89 (9): 1686–92. https://doi.org/10.1038/sj.bjc.6601340.
Garay-Sevilla, Ma Eugenia, Armando Rojas, Manuel Portero-Otin, and Jaime Uribarri. 2021. “Dietary AGEs as Exogenous Boosters of Inflammation.” Nutrients 13 (8): 2802. https://doi.org/10.3390/nu13082802.
Gencer, Baris, Luc Djousse, Omar T Al-Ramady, Nancy R. Cook, JoAnn E. Manson, and Christine M. Albert. 2021. “Effect of Long-Term Marine Omega-3 Fatty Acids Supplementation on the Risk of Atrial Fibrillation in Randomized Controlled Trials of Cardiovascular Outcomes: A Systematic Review and Meta-Analysis.” Circulation 144 (25): 1981–90. https://doi.org/10.1161/CIRCULATIONAHA.121.055654.
Ghawi, Sameer Khalil, Lisa Methven, and Keshavan Niranjan. 2013. “The Potential to Intensify Sulforaphane Formation in Cooked Broccoli (Brassica Oleracea Var. Italica) Using Mustard Seeds (Sinapis Alba).” Food Chemistry 138 (2): 1734–41. https://doi.org/10.1016/j.foodchem.2012.10.119.
Giordano, Antonio, and Giuseppina Tommonaro. 2019. “Curcumin and Cancer.” Nutrients 11 (10): 2376. https://doi.org/10.3390/nu11102376.
Goyal, Ankit, Vivek Sharma, Manvesh Kumar Sihag, A. K. Singh, Sumit Arora, and Latha Sabikhi. 2016. “Fortification of Dahi (Indian Yoghurt) with Omega-3 Fatty Acids Using Microencapsulated Flaxseed Oil Microcapsules.” Journal of Food Science and Technology 53 (5): 2422–33. https://doi.org/10.1007/s13197-016-2220-1.
“GRAS Notices.” n.d.a. Accessed August 25, 2025. https://www.hfpappexternal.fda.gov/scripts/fdcc/index.cfm?id=686&set=GRASNotices&utm_source=chatgpt.com.
———. n.d.b. Accessed October 23, 2025. https://hfpappexternal.fda.gov/scripts/fdcc/index.cfm?id=200&set=GRASNotices&utm_source=chatgpt.com.
Grivennikov, Sergei I., Florian R. Greten, and Michael Karin. 2010. “Immunity, Inflammation, and Cancer.” Cell 140 (6): 883–99. https://doi.org/10.1016/j.cell.2010.01.025.
Grosso, Giuseppe, Silvio Buscemi, Fabio Galvano, Antonio Mistretta, Stefano Marventano, Vanessa La Vela, Filippo Drago, Santi Gangi, Francesco Basile, and Antonio Biondi. 2013. “Mediterranean Diet and Cancer: Epidemiological Evidence and Mechanism of Selected Aspects.” BMC Surgery 13 Suppl 2 (Suppl 2): S14. https://doi.org/10.1186/1471-2482-13-S2-S14.
“Gut Myrosinases Convert Glucosinolates to Sulforaphane.” 2022. FoundMyFitness. https://www.foundmyfitness.com/stories/qtxhrl.
H. Lee Moffitt Cancer Center and Research Institute. 2020. “Randomized, Phase II Clinical Trial of Sulforaphane in Bladder Cancer Chemoprevention.” Clinical trial registration NCT03517995. clinicaltrials.gov. https://clinicaltrials.gov/study/NCT03517995.
Harris, William S., and Clemens Von Schacky. 2004. “The Omega-3 Index: A New Risk Factor for Death from Coronary Heart Disease?” Preventive Medicine 39 (1): 212–20. https://doi.org/10.1016/j.ypmed.2004.02.030.
Hatta, Muhammad Nur Adam, Ezanee Azlina Mohamad Hanif, Siok-Fong Chin, and Hui-min Neoh. 2021. “Pathogens and Carcinogenesis: A Review.” Biology 10 (6): 533. https://doi.org/10.3390/biology10060533.
Hegde, Mangala, Sosmitha Girisa, Bandari BharathwajChetty, Ravichandran Vishwa, and Ajaikumar B. Kunnumakkara. 2023. “Curcumin Formulations for Better Bioavailability: What We Learned from Clinical Trials Thus Far?” ACS Omega 8 (12): 10713–46. https://doi.org/10.1021/acsomega.2c07326.
Heiss, Elke, Christian Herhaus, Karin Klimo, Helmut Bartsch, and Clarissa Gerhäuser. 2001. “Nuclear Factor κB Is a Molecular Target for Sulforaphane-Mediated Anti-Inflammatory Mechanisms *.” Journal of Biological Chemistry 276 (34): 32008–15. https://doi.org/10.1074/jbc.M104794200.
Houghton, Christine A. 2019. “Sulforaphane: Its ‘Coming of Age’ as a Clinically Relevant Nutraceutical in the Prevention and Treatment of Chronic Disease.” Oxidative Medicine and Cellular Longevity 2019 (October): 2716870. https://doi.org/10.1155/2019/2716870.
———. 2023a. “The Rationale for Sulforaphane Favourably Influencing Gut Homeostasis and Gut–Organ Dysfunction: A Clinician’s Hypothesis.” International Journal of Molecular Sciences 24 (17): 13448. https://doi.org/10.3390/ijms241713448.
———. 2023b. “The Rationale for Sulforaphane Favourably Influencing Gut Homeostasis and Gut-Organ Dysfunction: A Clinician’s Hypothesis.” International Journal of Molecular Sciences 24 (17): 13448. https://doi.org/10.3390/ijms241713448.
“How Much Curcumin Is in My Turmeric Latte?” n.d. Amoda. Accessed August 12, 2025. https://amodatea.com/blogs/amoda-tea-blog/how-much-curcumin-is-in-my-turmeric-latte.
Howells, Lynne M., Chinenye O. O. Iwuji, Glen R. B. Irving, Shaun Barber, Harriet Walter, Zahirah Sidat, Nicola Griffin-Teall, et al. 2019a. “Curcumin Combined with FOLFOX Chemotherapy Is Safe and Tolerable in Patients with Metastatic Colorectal Cancer in a Randomized Phase IIa Trial.” The Journal of Nutrition 149 (7): 1133–39. https://doi.org/10.1093/jn/nxz029.
Howells, Lynne M, Chinenye O O Iwuji, Glen R B Irving, Shaun Barber, Harriet Walter, Zahirah Sidat, Nicola Griffin-Teall, et al. 2019b. “Curcumin Combined with FOLFOX Chemotherapy Is Safe and Tolerable in Patients with Metastatic Colorectal Cancer in a Randomized Phase IIa Trial.” The Journal of Nutrition 149 (7): 1133–39. https://doi.org/10.1093/jn/nxz029.
Hu, Chenqi, Aimee L. Eggler, Andrew D. Mesecar, and Richard B. van Breemen. 2011. “Modification of Keap1 Cysteine Residues by Sulforaphane.” Chemical Research in Toxicology 24 (4): 515–21. https://doi.org/10.1021/tx100389r.
“Invisible Numbers.” n.d. Accessed July 6, 2025. https://www.who.int/teams/noncommunicable-diseases/invisible-numbers.
Ishida, Masahiko, Masakazu Hara, Nobuko Fukino, Tomohiro Kakizaki, and Yasujiro Morimitsu. 2014. “Glucosinolate Metabolism, Functionality and Breeding for the Improvement of Brassicaceae Vegetables.” Breeding Science 64 (1): 48–59. https://doi.org/10.1270/jsbbs.64.48.
Javaid, Mustafa, Kadhim Kadhim, Bilal Bawamia, Timothy Cartlidge, Mohamed Farag, and Mohammad Alkhalil. 2024. “Bleeding Risk in Patients Receiving Omega‐3 Polyunsaturated Fatty Acids: A Systematic Review and Meta‐Analysis of Randomized Clinical Trials.” Journal of the American Heart Association 13 (10): e032390. https://doi.org/10.1161/JAHA.123.032390.
Judith, Passildas Jahanmohan, Bernadach Maureen, Pouget Mélanie, Kwiatkowski Fabrice, Vanpraagh-Doreau Isabelle, Dubray-Longeras Pascale, Abrial Catherine, et al. 2024. “Curcumin’s Effect in Advanced and Metastatic Breast Cancer Patients Treated with First or Second-Line Docetaxel: A Randomized Trial.” Health Science Reports 7 (9): e70052. https://doi.org/10.1002/hsr2.70052.
Kanai, Masashi, Atsushi Imaizumi, Yoshihiko Otsuka, Hiroki Sasaki, Momo Hashiguchi, Kazu Tsujiko, Shigemi Matsumoto, Hiroshi Ishiguro, and Tsutomu Chiba. 2012. “Dose-Escalation and Pharmacokinetic Study of Nanoparticle Curcumin, a Potential Anticancer Agent with Improved Bioavailability, in Healthy Human Volunteers.” Cancer Chemotherapy and Pharmacology 69 (1): 65–70. https://doi.org/10.1007/s00280-011-1673-1.
Kantor, Elizabeth D., Johanna W. Lampe, Ulrike Peters, Thomas L. Vaughan, and Emily White. 2014. “Long-Chain Omega-3 Polyunsaturated Fatty Acid Intake and Risk of Colorectal Cancer.” Nutrition and Cancer 66 (4): 716–27. https://doi.org/10.1080/01635581.2013.804101.
Karimi, Roxanne, Timothy P. Fitzgerald, and Nicholas S. Fisher. 2012. “A Quantitative Synthesis of Mercury in Commercial Seafood and Implications for Exposure in the United States.” Environmental Health Perspectives 120 (11): 1512–19. https://doi.org/10.1289/ehp.1205122.
Kim, Hee-Yong, Bill X. Huang, and Arthur A. Spector. 2022. “Molecular and Signaling Mechanisms for Docosahexaenoic Acid-Derived Neurodevelopment and Neuroprotection.” International Journal of Molecular Sciences 23 (9): 4635. https://doi.org/10.3390/ijms23094635.
Kim, Jae Kwang, and Sang Un Park. 2016. “Current Potential Health Benefits of Sulforaphane.” EXCLI Journal 15 (October): 571–77. https://doi.org/10.17179/excli2016-485.
Kocher, Alexa, Christina Schiborr, Dariush Behnam, and Jan Frank. 2015. “The Oral Bioavailability of Curcuminoids in Healthy Humans Is Markedly Enhanced by Micellar Solubilisation but Not Further Improved by Simultaneous Ingestion of Sesamin, Ferulic Acid, Naringenin and Xanthohumol.” Journal of Functional Foods 14 (April): 183–91. https://doi.org/10.1016/j.jff.2015.01.045.
Koelman, Liselot, Caue Egea Rodrigues, and Krasimira Aleksandrova. 2022. “Effects of Dietary Patterns on Biomarkers of Inflammation and Immune Responses: A Systematic Review and Meta-Analysis of Randomized Controlled Trials.” Advances in Nutrition 13 (1): 101–15. https://doi.org/10.1093/advances/nmab086.
Köhler, Anton, Essi Sarkkinen, Niina Tapola, Tarja Niskanen, and Inge Bruheim. 2015. “Bioavailability of Fatty Acids from Krill Oil, Krill Meal and Fish Oil in Healthy Subjects–a Randomized, Single-Dose, Cross-over Trial.” Lipids in Health and Disease 14 (1): 19. https://doi.org/10.1186/s12944-015-0015-4.
Konijeti, Gauree G., NaMee Kim, James D. Lewis, Shauna Groven, Anita Chandrasekaran, Sirisha Grandhe, Caroline Diamant, et al. 2017. “Efficacy of the Autoimmune Protocol Diet for Inflammatory Bowel Disease.” Inflammatory Bowel Diseases 23 (11): 2054–60. https://doi.org/10.1097/MIB.0000000000001221.
Kralik, Zlata, Gordana Kralik, Manuela Košević, Olivera Galović, Mirela Samardžić, Zlata Kralik, Gordana Kralik, Manuela Košević, Olivera Galović, and Mirela Samardžić. 2023. “Natural Multi-Enriched Eggs with n-3 Polyunsaturated Fatty Acids, Selenium, Vitamin E, and Lutein.” Animals 13 (2). https://doi.org/10.3390/ani13020321.
Kris-Etherton, Penny M., William S. Harris, and Lawrence J. Appel. 2002. “Fish Consumption, Fish Oil, Omega-3 Fatty Acids, and Cardiovascular Disease.” Circulation 106 (21): 2747–57. https://doi.org/10.1161/01.CIR.0000038493.65177.94.
Krupa, Kristina N., Kristina Fritz, and Mayur Parmar. 2025. “Omega-3 Fatty Acids.” In StatPearls. Treasure Island (FL): StatPearls Publishing. http://www.ncbi.nlm.nih.gov/books/NBK564314/.
Lao, Christopher D., Mack T. Ruffin IV, Daniel Normolle, Dennis D. Heath, Sandra I. Murray, Joanne M. Bailey, Martha E. Boggs, James Crowell, Cheryl L. Rock, and Dean E. Brenner. 2006. “Dose Escalation of a Curcuminoid Formulation.” BMC Complementary and Alternative Medicine 6 (10). https://doi.org/10.1186/1472-6882-6-10.
Lee, Young-Min, and Yoona Kim. 2024. “Is Curcumin Intake Really Effective for Chronic Inflammatory Metabolic Disease? A Review of Meta-Analyses of Randomized Controlled Trials.” Nutrients 16 (11): 1728. https://doi.org/10.3390/nu16111728.
Lee, Yu Geon, Do Hong Park, and Young Chan Chae. 2022. “Role of Mitochondrial Stress Response in Cancer Progression.” Cells 11 (5): 771. https://doi.org/10.3390/cells11050771.
Lefèvre, Sabine. 2024. “Is Cheese Good For A Mediterranean Diet? (Healthy Recipe Ideas).” Cheese Scientist. https://cheesescientist.com/science/cheese-for-mediterranean-diet/.
Liu, Danhui, Zhenyu Zhong, and Michael Karin. 2022. “NF-κB: A Double-Edged Sword Controlling Inflammation.” Biomedicines 10 (6): 1250. https://doi.org/10.3390/biomedicines10061250.
Liu, Ting, Lingyun Zhang, Donghyun Joo, and Shao-Cong Sun. 2017. “NF-κB Signaling in Inflammation.” Signal Transduction and Targeted Therapy 2 (1): 17023. https://doi.org/10.1038/sigtrans.2017.23.
Liu, Weidong, Yingjie Zhai, Xueyuan Heng, Feng Yuan Che, Wenjun Chen, Dezhong Sun, and Guangxi Zhai. 2016. “Oral Bioavailability of Curcumin: Problems and Advancements.” Journal of Drug Targeting 24 (8): 694–702. https://doi.org/10.3109/1061186X.2016.1157883.
Lopresti, Adrian L., Stephen J. Smith, Shavon Jackson-Michel, and Timothy Fairchild. 2022. “An Investigation into the Effects of a Curcumin Extract (Curcugen®) on Osteoarthritis Pain of the Knee: A Randomised, Double-Blind, Placebo-Controlled Study.” Nutrients 14 (1): 41. https://doi.org/10.3390/nu14010041.
lotan, yair. 2025. “A Randomized, Double-Blind, Placebo-Controlled Trial of Curcumin to Prevent Progression of Biopsy Proven, Low-Risk Localized Prostate Cancer Patients Undergoing Active Surveillance.” Clinical trial registration NCT03769766. clinicaltrials.gov. https://clinicaltrials.gov/study/NCT03769766.
Lubelska, Katarzyna, Małgorzata Milczarek, Karolina Modzelewska, Jolanta Krzysztoń-Russjan, Krzysztof Fronczyk, and Katarzyna Wiktorska. 2012a. “Interactions Between Drugs and Sulforaphane Modulate the Drug Metabolism Enzymatic System.” Pharmacological Reports: PR 64 (5): 1243–52. https://doi.org/10.1016/s1734-1140(12)70920-9.
———. 2012b. “Interactions Between Drugs and Sulforaphane Modulate the Drug Metabolism Enzymatic System.” Pharmacological Reports 64 (5): 1243–52. https://doi.org/10.1016/S1734-1140(12)70920-9.
Ma, Ziwei, Na Wang, Haibing He, and Xing Tang. 2019. “Pharmaceutical Strategies of Improving Oral Systemic Bioavailability of Curcumin for Clinical Application.” Journal of Controlled Release 316 (December): 359–80. https://doi.org/10.1016/j.jconrel.2019.10.053.
Manson, JoAnn E., Shari S. Bassuk, Julie E. Buring, and VITAL Research Group. 2020. “Principal Results of the VITamin D and OmegA-3 TriaL (VITAL) and Updated Meta-Analyses of Relevant Vitamin D Trials.” The Journal of Steroid Biochemistry and Molecular Biology 198 (April): 105522. https://doi.org/10.1016/j.jsbmb.2019.105522.
Marc_Grossman. 2016. “Three Ways to Improve Absorption of Turmeric or Curcumin.” Natural Eye Care Blog: News & Research on Vision. https://www.naturaleyecare.com/blog/improve-absorption-turmeric-curcumin/.
Marchi, Saverio, Emma Guilbaud, Stephen W. G. Tait, Takahiro Yamazaki, and Lorenzo Galluzzi. 2023. “Mitochondrial Control of Inflammation.” Nature Reviews Immunology 23 (3): 159–73. https://doi.org/10.1038/s41577-022-00760-x.
Mason, R. Preston, Peter Libby, and Deepak L. Bhatt. 2020. “Emerging Mechanisms of Cardiovascular Protection for the Omega-3 Fatty Acid Eicosapentaenoic Acid.” Arteriosclerosis, Thrombosis, and Vascular Biology 40 (5): 1135–47. https://doi.org/10.1161/ATVBAHA.119.313286.
Mecca, Marisabel, Marzia Sichetti, Martina Giuseffi, Eugenia Giglio, Claudia Sabato, Francesca Sanseverino, and Graziella Marino. 2024. “Synergic Role of Dietary Bioactive Compounds in Breast Cancer Chemoprevention and Combination Therapies.” Nutrients 16 (12): 1883. https://doi.org/10.3390/nu16121883.
MIAdmin. 2021. “Nrf-2 and Sulforaphane—What You Need to Know.” https://www.metagenicsinstitute.com/blogs/sulforaphane-nrf2-detoxification/.
Michels, Nathalie, Carola van Aart, Jens Morisse, Amy Mullee, and Inge Huybrechts. 2021. “Chronic Inflammation Towards Cancer Incidence: A Systematic Review and Meta-Analysis of Epidemiological Studies.” Critical Reviews in Oncology/Hematology 157 (January): 103177. https://doi.org/10.1016/j.critrevonc.2020.103177.
Miller, Elizabeth S., Camille G. Apple, Kolenkode B. Kannan, Zackary M. Funk, Jessica M. Plazas, Philip A. Efron, and Alicia M. Mohr. 2019. “Chronic Stress Induces Persistent Low-Grade Inflammation.” The American Journal of Surgery 218 (4): 677–83. https://doi.org/10.1016/j.amjsurg.2019.07.006.
Mocellin, Michel Carlos, Juliana de Aguiar Pastore e Silva, Carolina de Quadros Camargo, Maria Emília de Souza Fabre, Scheila Gevaerd, Katya Naliwaiko, Yara Maria Franco Moreno, Everson Araújo Nunes, and Erasmo Benicio Santos de Moraes Trindade. 2013. “Fish Oil Decreases C-Reactive Protein/Albumin Ratio Improving Nutritional Prognosis and Plasma Fatty Acid Profile in Colorectal Cancer Patients.” Lipids 48 (9): 879–88. https://doi.org/10.1007/s11745-013-3816-0.
Monllor-Tormos, Aitana, Alicia García-Vigara, Orly Morgan, Miguel-Ángel García-Pérez, Nicolás Mendoza, Juan J. Tarín, and Antonio Cano. 2023. “Mediterranean Diet for Cancer Prevention and Survivorship.” Maturitas 178 (December). https://doi.org/10.1016/j.maturitas.2023.107841.
Mukhopadhyay, Asok, Sanjeev Banerjee, Lewis Joe Stafford, Chunzhi Xia, Mingyao Liu, and Bharat B. Aggarwal. 2002. “Curcumin-Induced Suppression of Cell Proliferation Correlates with down-Regulation of Cyclin D1 Expression and CDK4-Mediated Retinoblastoma Protein Phosphorylation.” Oncogene 21 (57): 8852–61. https://doi.org/10.1038/sj.onc.1206048.
Munday, Rex, Paulette Mhawech-Fauceglia, Christine M. Munday, Joseph D. Paonessa, Li Tang, John S. Munday, Carolyn Lister, et al. 2008. “Inhibition of Urinary Bladder Carcinogenesis by Broccoli Sprouts.” Cancer Research 68 (5): 1593–1600. https://doi.org/10.1158/0008-5472.CAN-07-5009.
Murphy, Rachel A., Marina Mourtzakis, Quincy S. C. Chu, Vickie E. Baracos, Tony Reiman, and Vera C. Mazurak. 2011. “Supplementation with Fish Oil Increases First-Line Chemotherapy Efficacy in Patients with Advanced Nonsmall Cell Lung Cancer.” Cancer 117 (16): 3774–80. https://doi.org/10.1002/cncr.25933.
Muscaritoli, Maurizio, Jann Arends, Patrick Bachmann, Vickie Baracos, Nicole Barthelemy, Hartmut Bertz, Federico Bozzetti, et al. 2021. “ESPEN Practical Guideline: Clinical Nutrition in Cancer.” Clinical Nutrition 40 (5): 2898–2913. https://doi.org/10.1016/j.clnu.2021.02.005.
Myzak, Melinda C., P. Andrew Karplus, Fung-Lung Chung, and Roderick H. Dashwood. 2004. “A Novel Mechanism of Chemoprotection by Sulforaphane: Inhibition of Histone Deacetylase.” Cancer Research 64 (16): 5767–74. https://doi.org/10.1158/0008-5472.CAN-04-1326.
“NIH-AARP Diet and Health Study – NCI.” 2022. {cgvArticle}. https://dceg.cancer.gov/research/who-we-study/nih-aarp-diet-health-study.
“Non Communicable Diseases.” n.d. Accessed July 6, 2025. https://www.who.int/news-room/fact-sheets/detail/noncommunicable-diseases.
O’Keefe, Stephen J. D., Jia V. Li, Leo Lahti, Junhai Ou, Franck Carbonero, Khaled Mohammed, Joram M. Posma, et al. 2015. “Fat, Fibre and Cancer Risk in African Americans and Rural Africans.” Nature Communications 6 (1): 6342. https://doi.org/10.1038/ncomms7342.
“Office of Dietary Supplements – Omega-3 Fatty Acids.” n.d.a. Accessed August 28, 2025. https://ods.od.nih.gov/factsheets/Omega3FattyAcids-HealthProfessional/.
———. n.d.b. Accessed October 6, 2025. https://ods.od.nih.gov/factsheets/Omega3FattyAcids-HealthProfessional/.
Oloyede, Omobolanle O., Carol Wagstaff, and Lisa Methven. 2021. “The Impact of Domestic Cooking Methods on Myrosinase Stability, Glucosinolates and Their Hydrolysis Products in Different Cabbage (Brassica Oleracea) Accessions.” Foods 10 (12): 2908. https://doi.org/10.3390/foods10122908.
“Omega-3 Supplements: What You Need To Know.” n.d. NCCIH. Accessed October 23, 2025. https://www.nccih.nih.gov/health/omega3-supplements-what-you-need-to-know.
Organisation des Nations Unies pour l’alimentation et l’agriculture, and Organisation mondiale de la santé, eds. 2010a. Fats and Fatty Acids in Human Nutrition: Report of an Expert Consultation 10-14 November 2008, Geneva. FAO Food and Nutrition Paper 91. Rome: FAO.
———, eds. 2010b. Fats and Fatty Acids in Human Nutrition: Report of an Expert Consultation 10-14 November 2008, Geneva. FAO Food and Nutrition Paper 91. Rome: FAO.
Otoo, Raymond A., and Antiño R. Allen. 2023. “Sulforaphane’s Multifaceted Potential: From Neuroprotection to Anticancer Action.” Molecules 28 (19): 6902. https://doi.org/10.3390/molecules28196902.
Pa, Egner, Chen Jg, Zarth At, Ng Dk, Wang Jb, Kensler Kh, Jacobson Lp, et al. 2014. “Rapid and Sustainable Detoxication of Airborne Pollutants by Broccoli Sprout Beverage: Results of a Randomized Clinical Trial in China.” Cancer Prevention Research (Philadelphia, Pa.) 7 (8). https://doi.org/10.1158/1940-6207.CAPR-14-0103.
Palliyaguru, Dushani L., Jian-Min Yuan, Thomas W. Kensler, and Jed W. Fahey. 2018. “Isothiocyanates: Translating the Power of Plants to People.” Molecular Nutrition & Food Research 62 (18): 1700965. https://doi.org/10.1002/mnfr.201700965.
Panahi, Yunes, Nahid Khalili, Ebrahim Sahebi, Soha Namazi, Željko Reiner, Muhammed Majeed, and Amirhossein Sahebkar. 2017. “Curcuminoids Modify Lipid Profile in Type 2 Diabetes Mellitus: A Randomized Controlled Trial.” Complementary Therapies in Medicine 33 (August): 1–5. https://doi.org/10.1016/j.ctim.2017.05.006.
Pandey, Ashish, Fozia Kamran, Manisha Choudhury, Li Li, Mohammad Shafiur Rahman, and Malik Altaf Hussain. 2025. “Omega-3 Fatty Acid Fortification of Plant-Based Beverages to Enhance Their Nutritional Profile.” Foods 14 (9): 1602. https://doi.org/10.3390/foods14091602.
Paśko, Paweł, Paweł Zagrodzki, Krzysztof Okoń, Ewelina Prochownik, Mirosław Krośniak, and Agnieszka Galanty. 2022. “Broccoli Sprouts and Their Influence on Thyroid Function in Different In Vitro and In Vivo Models.” Plants 11 (20): 2750. https://doi.org/10.3390/plants11202750.
Patterson, E., R. Wall, G. F. Fitzgerald, R. P. Ross, and C. Stanton. 2012. “Health Implications of High Dietary Omega-6 Polyunsaturated Fatty Acids.” Journal of Nutrition and Metabolism 2012: 539426. https://doi.org/10.1155/2012/539426.
PDQ Integrative, Alternative, and Complementary Therapies Editorial Board. 2002. “Curcumin (Curcuma, Turmeric) and Cancer (PDQ®): Patient Version.” In PDQ Cancer Information Summaries. Bethesda (MD): National Cancer Institute (US). http://www.ncbi.nlm.nih.gov/books/NBK578436/.
Pogorzelska, A., M. Świtalska, J. Wietrzyk, M. Mazur, M. Milczarek, K. Medyńska, and K. Wiktorska. 2024. “Antitumor and Antimetastatic Effects of Dietary Sulforaphane in a Triple-Negative Breast Cancer Models.” Scientific Reports 14 (1): 16016. https://doi.org/10.1038/s41598-024-65455-w.
Priyadarsini, K. Indira, Dilip K. Maity, G. H. Naik, M. Sudheer Kumar, M. K. Unnikrishnan, J. G. Satav, and Hari Mohan. 2003. “Role of Phenolic O-H and Methylene Hydrogen on the Free Radical Reactions and Antioxidant Activity of Curcumin.” Free Radical Biology and Medicine 35 (5): 475–84. https://doi.org/10.1016/S0891-5849(03)00325-3.
Program, Human Foods. 2024. “Advice about Eating Fish.” FDA, September. https://www.fda.gov/food/consumers/advice-about-eating-fish.
Program, National Toxicology. 2021. “Asbestos.” In 15th Report on Carcinogens [Internet]. National Toxicology Program. https://www.ncbi.nlm.nih.gov/books/NBK590791/.
“Prostaphane.” n.d. Prostaphane. Accessed November 17, 2025. https://www.prostaphane.com/prostaphane/.
“Prostate Cancer Prevention Trial (PCPT): Questions and Answers – NCI.” n.d. Accessed September 22, 2025. https://www.cancer.gov/types/prostate/research/prostate-cancer-prevention-trial-qa.
Qiu, Peiyu, Shuli Man, Jing Li, Jing Liu, Liming Zhang, Peng Yu, and Wenyuan Gao. 2016. “Overdose Intake of Curcumin Initiates the Unbalanced State of Bodies.” Journal of Agricultural and Food Chemistry 64 (13): 2765–71. https://doi.org/10.1021/acs.jafc.6b00053.
“Quantitation of Alpha-Linolenic Acid Elongation to Eicosapentaenoic and Docosahexaenoic Acid as Affected by the Ratio of N6/N3 Fatty Acids Nutrition & Metabolism Full Text.” n.d. Accessed August 28, 2025. https://nutritionandmetabolism.biomedcentral.com/articles/10.1186/1743-7075-6-8.
Rajaram, Sujatha. 2014. “Health Benefits of Plant-Derived α-Linolenic Acid.” The American Journal of Clinical Nutrition 100 (suppl_1): 443S–448S. https://doi.org/10.3945/ajcn.113.071514.
Rajendran, Peramaiyan, Ya-Fang Chen, Yu-Feng Chen, Li-Chin Chung, Shanmugam Tamilselvi, Chia-Yao Shen, Cecilia Hsuan Day, et al. 2018. “The Multifaceted Link Between Inflammation and Human Diseases.” Journal of Cellular Physiology 233 (9): 6458–71. https://doi.org/10.1002/jcp.26479.
Restivo, Jenette. 2024. “Omega-3 Foods: Incorporating Healthy Fats into Your Diet.” Harvard Health. https://www.health.harvard.edu/nutrition/omega-3-foods-incorporating-healthy-fats-into-your-diet.
Satokari, Reetta. 2020. “High Intake of Sugar and the Balance Between Pro- and Anti-Inflammatory Gut Bacteria.” Nutrients 12 (5): 1348. https://doi.org/10.3390/nu12051348.
Schuchardt, Jan Philipp, Inga Schneider, Henrike Meyer, Juliane Neubronner, Clemens von Schacky, and Andreas Hahn. 2011. “Incorporation of EPA and DHA into Plasma Phospholipids in Response to Different Omega-3 Fatty Acid Formulations – a Comparative Bioavailability Study of Fish Oil Vs. Krill Oil.” Lipids in Health and Disease 10 (1): 145. https://doi.org/10.1186/1476-511X-10-145.
Schwingshackl, L., and G. Hoffmann. 2014. “Mediterranean Dietary Pattern, Inflammation and Endothelial Function: A Systematic Review and Meta-Analysis of Intervention Trials.” Nutrition, Metabolism and Cardiovascular Diseases 24 (9): 929–39. https://doi.org/10.1016/j.numecd.2014.03.003.
Shahcheraghi, Seyed Hossein, Fateme Salemi, Niloufar Peirovi, Jamshid Ayatollahi, Waqas Alam, Haroon Khan, and Luciano Saso. 2022. “Nrf2 Regulation by Curcumin: Molecular Aspects for Therapeutic Prospects.” Molecules 27 (1): 167. https://doi.org/10.3390/molecules27010167.
Shapiro, A, L Wade, and K Stephenson. n.d. “Human Metabolism and Excretion of Cancer Chemoprotective Glucosinolates and Isothiocyanates of Cruciferous.”
Shoba, Guido, David Joy, Thangam Joseph, M. Majeed, R. Rajendran, and P. S. S. R. Srinivas. 2007. “Influence of Piperine on the Pharmacokinetics of Curcumin in Animals and Human Volunteers.” Planta Medica 64 (January): 353–56. https://doi.org/10.1055/s-2006-957450.
Simopoulos, A. P. 2002. “The Importance of the Ratio of Omega-6/Omega-3 Essential Fatty Acids.” Biomedicine & Pharmacotherapy 56 (8): 365–79. https://doi.org/10.1016/S0753-3322(02)00253-6.
“Singapore Chinese Health Study (SCHS).” n.d. Saw Swee Hock School of Public Health. Accessed September 22, 2025. https://sph.nus.edu.sg/research/cohort-schs/.
Singh, Ajita V., Dong Xiao, Karen L. Lew, Rajiv Dhir, and Shivendra V. Singh. 2004. “Sulforaphane Induces Caspase-Mediated Apoptosis in Cultured PC-3 Human Prostate Cancer Cells and Retards Growth of PC-3 Xenografts in Vivo.” Carcinogenesis 25 (1): 83–90. https://doi.org/10.1093/carcin/bgg178.
Singh, Nitin, Deepak Baby, Jagadish Prasad Rajguru, Pankaj B Patil, Savita S Thakkannavar, and Veena Bhojaraj Pujari. 2019. “Inflammation and Cancer.” Annals of African Medicine 18 (3): 121–26. https://doi.org/10.4103/aam.aam_56_18.
Skulas-Ray, Ann C., Peter W. F. Wilson, William S. Harris, Eliot A. Brinton, Penny M. Kris-Etherton, Chesney K. Richter, Terry A. Jacobson, et al. 2019. “Omega-3 Fatty Acids for the Management of Hypertriglyceridemia: A Science Advisory From the American Heart Association.” Circulation 140 (12): e673–91. https://doi.org/10.1161/CIR.0000000000000709.
Sohrab, Sayed Sartaj, Riya Raj, Amka Nagar, Susan Hawthorne, Ana Cláudia Paiva-Santos, Mohammad Amjad Kamal, Mai M. El-Daly, Esam I. Azhar, and Ankur Sharma. 2023. “Chronic Inflammation’s Transformation to Cancer: A Nanotherapeutic Paradigm.” Molecules (Basel, Switzerland) 28 (11): 4413. https://doi.org/10.3390/molecules28114413.
Stanimirovic, Julijana, Jelena Radovanovic, Katarina Banjac, Milan Obradovic, Magbubah Essack, Sonja Zafirovic, Zoran Gluvic, Takashi Gojobori, and Esma R. Isenovic. 2022. “Role of C-Reactive Protein in Diabetic Inflammation.” Mediators of Inflammation 2022 (1): 3706508. https://doi.org/10.1155/2022/3706508.
“Sulforaphane – an Overview ScienceDirect Topics.” n.d. Accessed November 18, 2025. https://www.sciencedirect.com/topics/pharmacology-toxicology-and-pharmaceutical-science/sulforaphane.
“Sulforaphane: Benefits, Side Effects, and Food Sources.” 2019. Healthline. https://www.healthline.com/nutrition/sulforaphane.
Surette, Marc E. 2008. “The Science Behind Dietary Omega-3 Fatty Acids.” CMAJ 178 (2): 177–80. https://doi.org/10.1503/cmaj.071356.
Tayyem, Reema F., Dennis D. Heath, Wael K. Al-Delaimy, and Cheryl L. Rock. 2006. “Curcumin Content of Turmeric and Curry Powders.” Nutrition and Cancer 55 (2): 126–31. https://doi.org/10.1207/s15327914nc5502_2.
“Testing the Effect of Sulforaphane, a Compound Naturally Found in Cruciferous Vegetables, on Preventing Melanoma in Patients With a Prior History of Melanoma.” n.d. Accessed November 14, 2025. https://prevention.cancer.gov/clinical-trials/clinical-trials-search/nct07040280.
Thanh, Lam Phuoc, Juan J. Loor, Duong Tran Tuyet Mai, and Tran Thi Thuy Hang. 2023. “Effect of Fish Oil and Linseed Oil on Intake, Milk Yield and Milk Fatty Acid Profile in Goats.” Animals : An Open Access Journal from MDPI 13 (13): 2174. https://doi.org/10.3390/ani13132174.
“Thorne Curcumin Phytosome (Formerly Meriva).” n.d. Accessed August 5, 2025. https://store.mayoclinic.com/thorne-meriva-500-sf.html.
“Tolerable Upper Intake Level of EPA, DHA and DPA EFSA.” 2012. https://www.efsa.europa.eu/en/efsajournal/pub/2815.
Tripathi, Siddhant, Yashika Sharma, and Dileep Kumar. 2025. “Unveiling the Link Between Chronic Inflammation and Cancer.” Metabolism Open 25 (March): 100347. https://doi.org/10.1016/j.metop.2025.100347.
University of Kansas Medical Center. 2023. “Omega 3 Fatty Acids in Colorectal Cancer (CRC) Prevention in Patients With Lynch Syndrome (COLYNE).” Clinical trial registration NCT03831698. clinicaltrials.gov. https://clinicaltrials.gov/study/NCT03831698.
Valizadeh Kiamahalleh, Mohammad, Ghasem Najafpour-Darzi, Mostafa Rahimnejad, Ali Akbar Moghadamnia, and Meisam Valizadeh Kiamahalleh. 2016. “High Performance Curcumin Subcritical Water Extraction from Turmeric (Curcuma Longa L.).” Journal of Chromatography B 1022 (June): 191–98. https://doi.org/10.1016/j.jchromb.2016.04.021.
“Vitamin A.” 2012. In LiverTox: Clinical and Research Information on Drug-Induced Liver Injury. Bethesda (MD): National Institute of Diabetes; Digestive; Kidney Diseases. http://www.ncbi.nlm.nih.gov/books/NBK548165/.
Wang, Haijun, Ke Zhang, Jia Liu, Jie Yang, Yidan Tian, Chen Yang, Yushan Li, Minglong Shao, Wei Su, and Na Song. 2021. “Curcumin Regulates Cancer Progression: Focus on ncRNAs and Molecular Signaling Pathways.” Frontiers in Oncology 11 (April). https://doi.org/10.3389/fonc.2021.660712.
Waure, Chiara de, Carlotta Bertola, Gaia Baccarini, Manuela Chiavarini, and Cesare Mancuso. 2023. “Exploring the Contribution of Curcumin to Cancer Therapy: A Systematic Review of Randomized Controlled Trials.” Pharmaceutics 15 (4): 1275. https://doi.org/10.3390/pharmaceutics15041275.
Wen, Yalei, Yingjie Zhu, Caishi Zhang, Xiao Yang, Yuchen Gao, Mei Li, Hongyan Yang, Tongzheng Liu, and Hui Tang. 2022. “Chronic Inflammation, Cancer Development and Immunotherapy.” Frontiers in Pharmacology 13 (October). https://doi.org/10.3389/fphar.2022.1040163.
White, Emily, Ruth E. Patterson, Alan R. Kristal, Mark Thornquist, Irena King, Ann L. Shattuck, Ilonka Evans, Jessie Satia-Abouta, Alyson J. Littman, and John D. Potter. 2004. “VITamins And Lifestyle Cohort Study: Study Design and Characteristics of Supplement Users.” American Journal of Epidemiology 159 (1): 83–93. https://doi.org/10.1093/aje/kwh010.
Wu, Xian, Lin Zhong, Yuehong Hu, Lingying Ruan, Kaiyu Zhou, Hanmin Liu, and Lina Chen. 2025. “Sedentary Behavior Is Associated with Systemic Immune-Inflammation Index and Systemic Inflammation Response Index Levels: A Cross-Sectional Analysis of the NHANES 2011–2018.” Frontiers in Public Health 13 (March). https://doi.org/10.3389/fpubh.2025.1431065.
www.prohealth.com. n.d. “Liposomal Sulforaphane: A Broccoli-Based Antioxidant with Enhanced Absorption That Supports Health With Age.” ProHealth.com. Accessed November 17, 2025. https://www.prohealth.com/blogs/control-how-you-age/liposomal-sulforaphane-a-broccoli-based-antioxidant-with-enhanced-absorption-that-supports-health-with-age.
Yagishita, Yoko, Jed W. Fahey, Albena T. Dinkova-Kostova, and Thomas W. Kensler. 2019. “Broccoli or Sulforaphane: Is It the Source or Dose That Matters?” Molecules 24 (19): 3593. https://doi.org/10.3390/molecules24193593.
Yanaka, Akinori, Jed W. Fahey, Atsushi Fukumoto, Mari Nakayama, Souta Inoue, Songhua Zhang, Masafumi Tauchi, Hideo Suzuki, Ichinosuke Hyodo, and Masayuki Yamamoto. 2009a. “Dietary Sulforaphane-Rich Broccoli Sprouts Reduce Colonization and Attenuate Gastritis in Helicobacter Pylori-Infected Mice and Humans.” Cancer Prevention Research (Philadelphia, Pa.) 2 (4): 353–60. https://doi.org/10.1158/1940-6207.CAPR-08-0192.
———. 2009b. “Dietary Sulforaphane-Rich Broccoli Sprouts Reduce Colonization and Attenuate Gastritis in Helicobacter Pylori–Infected Mice and Humans.” Cancer Prevention Research 2 (4): 353–60. https://doi.org/10.1158/1940-6207.CAPR-08-0192.
Yu, Weihua, Yongmei Tu, Zi Long, Jiangzheng Liu, Deqin Kong, Jie Peng, Hao Wu, et al. 2022. “Reactive Oxygen Species Bridge the Gap Between Chronic Inflammation and Tumor Development.” Oxidative Medicine and Cellular Longevity 2022 (June): 2606928. https://doi.org/10.1155/2022/2606928.
Yuan, Jian-Min, Thomas W. Kensler, Sanja Dacic, Douglas J. Hartman, Renwei Wang, Paula A. Balogh, Pamela Sufka, et al. 2025. “Randomized Phase II Clinical Trial of Sulforaphane in Former Smokers at High Risk for Lung Cancer.” Cancer Prevention Research (Philadelphia, Pa.) 18 (6): 335–45. https://doi.org/10.1158/1940-6207.CAPR-24-0386.
Yuanfeng, Wu, Lv Chengzhi, Zou Ligen, Sun Juan, Song Xinjie, Zhang Yao, and Mao Jianwei. 2021. “Approaches for Enhancing the Stability and Formation of Sulforaphane.” Food Chemistry 345 (May): 128771. https://doi.org/10.1016/j.foodchem.2020.128771.
Zhang, Haoyan, Sicheng Wang, Peng Wu, Zanmin Hu, Yani Chen, Yupeng Guan, and Jun Pang. 2025. “Revealing the Landscape of Targeting Mitochondrial Functions and Behaviors to Overcome Cancer Chemoresistance.” Journal of the National Cancer Center, June. https://doi.org/10.1016/j.jncc.2025.02.007.
Zhang, Y., P. Talalay, C. G. Cho, and G. H. Posner. 1992. “A Major Inducer of Anticarcinogenic Protective Enzymes from Broccoli: Isolation and Elucidation of Structure.” Proceedings of the National Academy of Sciences of the United States of America 89 (6): 2399–2403. https://doi.org/10.1073/pnas.89.6.2399.
Zheng, Dantong, Chongxing Huang, Haohe Huang, Yuan Zhao, Muhammad Rafi Ullah Khan, Hui Zhao, and Lijie Huang. 2020. “Antibacterial Mechanism of Curcumin: A Review.” Chemistry & Biodiversity 17 (8): e2000171. https://doi.org/10.1002/cbdv.202000171.
Zielińska, Aleksandra, Henrique Alves, Vânia Marques, Alessandra Durazzo, Massimo Lucarini, Thais F. Alves, Margreet Morsink, et al. 2020. “Properties, Extraction Methods, and Delivery Systems for Curcumin as a Natural Source of Beneficial Health Effects.” Medicina 56 (7): 336. https://doi.org/10.3390/medicina56070336.
Zotova, Natalya, Yulia Zhuravleva, Valeriy Chereshnev, and Evgenii Gusev. 2023. “Acute and Chronic Systemic Inflammation: Features and Differences in the Pathogenesis, and Integral Criteria for Verification and Differentiation.” International Journal of Molecular Sciences 24 (2): 1144. https://doi.org/10.3390/ijms24021144.
Zou, Sailan, Qiyu Tong, Bowen Liu, Wei Huang, Yan Tian, and Xianghui Fu. 2020. “Targeting STAT3 in Cancer Immunotherapy.” Molecular Cancer 19 (1): 145. https://doi.org/10.1186/s12943-020-01258-7.
Natalya Zotova et al., “Acute and Chronic Systemic Inflammation: Features and Differences in the Pathogenesis, and Integral Criteria for Verification and Differentiation,” International Journal of Molecular Sciences 24, no. 2 (January 2023): 1144, https://doi.org/10.3390/ijms24021144.↩︎
Peramaiyan Rajendran et al., “The Multifaceted Link between Inflammation and Human Diseases,” Journal of Cellular Physiology 233, no. 9 (2018): 6458–71, https://doi.org/10.1002/jcp.26479.↩︎
Sayed Sartaj Sohrab et al., “Chronic Inflammation’s Transformation to Cancer: A Nanotherapeutic Paradigm,” Molecules (Basel, Switzerland) 28, no. 11 (May 29, 2023): 4413, https://doi.org/10.3390/molecules28114413.↩︎
“Non Communicable Diseases,” accessed July 6, 2025, https://www.who.int/news-room/fact-sheets/detail/noncommunicable-diseases.
↩︎“Invisible Numbers,” accessed July 6, 2025, https://www.who.int/teams/noncommunicable-diseases/invisible-numbers.↩︎
“Cancer,” accessed July 6, 2025, https://www.who.int/news-room/fact-sheets/detail/cancer.↩︎
“Cancer Today,” accessed July 6, 2025, https://gco.iarc.who.int/today/.↩︎
Hannah Arem and Erikka Loftfield, “Cancer Epidemiology: A Survey of Modifiable Risk Factors for Prevention and Survivorship,” American Journal of Lifestyle Medicine 12, no. 3 (2017): 200–210, https://doi.org/10.1177/1559827617700600.↩︎
Danhui Liu et al., “NF-κB: A Double-Edged Sword Controlling Inflammation,” Biomedicines 10, no. 6 (2022): 6, https://doi.org/10.3390/biomedicines10061250.↩︎
Ting Liu et al., “NF-κB Signaling in Inflammation,” Signal Transduction and Targeted Therapy 2, no. 1 (2017): 17023, https://doi.org/10.1038/sigtrans.2017.23.↩︎
Ashley Beam et al., “Effect of Diet and Dietary Components on the Composition of the Gut Microbiota,” Nutrients 13, no. 8 (2021): 2795, https://doi.org/10.3390/nu13082795.↩︎
Reetta Satokari, “High Intake of Sugar and the Balance between Pro- and Anti-Inflammatory Gut Bacteria,” Nutrients 12, no. 5 (2020): 5, https://doi.org/10.3390/nu12051348.↩︎
Ma Eugenia Garay-Sevilla et al., “Dietary AGEs as Exogenous Boosters of Inflammation,” Nutrients 13, no. 8 (2021): 8, https://doi.org/10.3390/nu13082802.↩︎
Xian Wu et al., “Sedentary Behavior Is Associated with Systemic Immune-Inflammation Index and Systemic Inflammation Response Index Levels: A Cross-Sectional Analysis of the NHANES 2011–2018,” Frontiers in Public Health 13 (March 2025), https://doi.org/10.3389/fpubh.2025.1431065.↩︎
Massimo Fioranelli et al., “Stress and Inflammation in Coronary Artery Disease: A Review Psychoneuroendocrineimmunology-Based,” Frontiers in Immunology 9 (September 2018), https://doi.org/10.3389/fimmu.2018.02031.↩︎
Julijana Stanimirovic et al., “Role of C-Reactive Protein in Diabetic Inflammation,” Mediators of Inflammation 2022, no. 1 (2022): 3706508, https://doi.org/10.1155/2022/3706508.↩︎
Elizabeth S. Miller et al., “Chronic Stress Induces Persistent Low-Grade Inflammation,” The American Journal of Surgery 218, no. 4 (2019): 677–83, https://doi.org/10.1016/j.amjsurg.2019.07.006.↩︎
Siddhant Tripathi et al., “Unveiling the Link between Chronic Inflammation and Cancer,” Metabolism Open 25 (March 2025): 100347, https://doi.org/10.1016/j.metop.2025.100347.↩︎
Nathalie Michels et al., “Chronic Inflammation towards Cancer Incidence: A Systematic Review and Meta-Analysis of Epidemiological Studies,” Critical Reviews in Oncology/Hematology 157 (January 2021): 103177, https://doi.org/10.1016/j.critrevonc.2020.103177.↩︎
Muhammad Nur Adam Hatta et al., “Pathogens and Carcinogenesis: A Review,” Biology 10, no. 6 (2021): 6, https://doi.org/10.3390/biology10060533.↩︎
Sergei I. Grivennikov et al., “Immunity, Inflammation, and Cancer,” Cell 140, no. 6 (2010): 883–99, https://doi.org/10.1016/j.cell.2010.01.025.↩︎
Nitin Singh et al., “Inflammation and Cancer,” Annals of African Medicine 18, no. 3 (2019): 121–26, https://doi.org/10.4103/aam.aam_56_18.↩︎
National Toxicology Program, “Asbestos,” in 15th Report on Carcinogens [Internet] (National Toxicology Program, 2021), https://www.ncbi.nlm.nih.gov/books/NBK590791/.↩︎
Paul J. A. Borm et al., “An Updated Review of the Genotoxicity of Respirable Crystalline Silica,” Particle and Fibre Toxicology 15, no. 1 (2018): 23, https://doi.org/10.1186/s12989-018-0259-z.↩︎
Yalei Wen et al., “Chronic Inflammation, Cancer Development and Immunotherapy,” Frontiers in Pharmacology 13 (October 2022), https://doi.org/10.3389/fphar.2022.1040163.↩︎
Yaning Cao et al., “NF-κB Signaling Pathway in Tumor Microenvironment,” Frontiers in Immunology 15 (October 2024), https://doi.org/10.3389/fimmu.2024.1476030.↩︎
Weihua Yu et al., “Reactive Oxygen Species Bridge the Gap between Chronic Inflammation and Tumor Development,” Oxidative Medicine and Cellular Longevity 2022 (June 2022): 2606928, https://doi.org/10.1155/2022/2606928.↩︎
Saverio Marchi et al., “Mitochondrial Control of Inflammation,” Nature Reviews Immunology 23, no. 3 (2023): 159–73, https://doi.org/10.1038/s41577-022-00760-x.↩︎
Haoyan Zhang et al., “Revealing the Landscape of Targeting Mitochondrial Functions and Behaviors to Overcome Cancer Chemoresistance,” Journal of the National Cancer Center, ahead of print, June 8, 2025, https://doi.org/10.1016/j.jncc.2025.02.007.↩︎
Yu Geon Lee et al., “Role of Mitochondrial Stress Response in Cancer Progression,” Cells 11, no. 5 (2022): 771, https://doi.org/10.3390/cells11050771.↩︎
Cornelia Braicu et al., “A Comprehensive Review on MAPK: A Promising Therapeutic Target in Cancer,” Cancers 11, no. 10 (2019): 10, https://doi.org/10.3390/cancers11101618.↩︎
Sailan Zou et al., “Targeting STAT3 in Cancer Immunotherapy,” Molecular Cancer 19, no. 1 (2020): 145, https://doi.org/10.1186/s12943-020-01258-7.↩︎
Dolores Aguilar-Cazares et al., “Contribution of Angiogenesis to Inflammation and Cancer,” Frontiers in Oncology 9 (December 2019), https://doi.org/10.3389/fonc.2019.01399.↩︎
Laura S. Angelo and Razelle Kurzrock, “Vascular Endothelial Growth Factor and Its Relationship to Inflammatory Mediators,” Clinical Cancer Research 13, no. 10 (2007): 2825–30, https://doi.org/10.1158/1078-0432.CCR-06-2416.↩︎
R. Badmanaban et al., “Turmeric: A Holistic Solution for Biochemical Malfunction,” Research Journal of Pharmacy and Technology 14, no. 10 (2021): 5540–50, https://doi.org/10.52711/0974-360X.2021.00966.↩︎
Dantong Zheng et al., “Antibacterial Mechanism of Curcumin: A Review,” Chemistry & Biodiversity 17, no. 8 (2020): e2000171, https://doi.org/10.1002/cbdv.202000171.↩︎
Tuba Ak and İlhami Gülçin, “Antioxidant and Radical Scavenging Properties of Curcumin,” Chemico-Biological Interactions 174, no. 1 (2008): 27–37, https://doi.org/10.1016/j.cbi.2008.05.003.↩︎
Seyed Hossein Shahcheraghi et al., “Nrf2 Regulation by Curcumin: Molecular Aspects for Therapeutic Prospects,” Molecules 27, no. 1 (2022): 1, https://doi.org/10.3390/molecules27010167.↩︎
Bharat B. Aggarwal and Kuzhuvelil B. Harikumar, “Potential Therapeutic Effects of Curcumin, the Anti-Inflammatory Agent, against Neurodegenerative, Cardiovascular, Pulmonary, Metabolic, Autoimmune and Neoplastic Diseases,” The International Journal of Biochemistry & Cell Biology, Directed Issue: Epigenetics and Disease, vol. 41, no. 1 (2009): 40–59, https://doi.org/10.1016/j.biocel.2008.06.010.↩︎
Antonio Giordano and Giuseppina Tommonaro, “Curcumin and Cancer,” Nutrients 11, no. 10 (2019): 10, https://doi.org/10.3390/nu11102376.↩︎
Asok Mukhopadhyay et al., “Curcumin-Induced Suppression of Cell Proliferation Correlates with down-Regulation of Cyclin D1 Expression and CDK4-Mediated Retinoblastoma Protein Phosphorylation,” Oncogene 21, no. 57 (2002): 8852–61, https://doi.org/10.1038/sj.onc.1206048.↩︎
Sabrina Bimonte et al., “Dissecting the Role of Curcumin in Tumour Growth and Angiogenesis in Mouse Model of Human Breast Cancer,” BioMed Research International 2015 (2015): 878134, https://doi.org/10.1155/2015/878134.↩︎
Lynne M. Howells et al., “Curcumin Combined with FOLFOX Chemotherapy Is Safe and Tolerable in Patients with Metastatic Colorectal Cancer in a Randomized Phase IIa Trial,” The Journal of Nutrition 149, no. 7 (2019): 1133–39, https://doi.org/10.1093/jn/nxz029.↩︎
Robert E. Carroll et al., “Phase IIA Clinical Trial of Curcumin for the Prevention of Colorectal Neoplasia,” Cancer Prevention Research (Philadelphia, Pa.) 4, no. 3 (2011): 354–64, https://doi.org/10.1158/1940-6207.CAPR-10-0098.↩︎
Marcia Cruz-Correa et al., “Combination Treatment with Curcumin and Quercetin of Adenomas in Familial Adenomatous Polyposis,” Clinical Gastroenterology and Hepatology: The Official Clinical Practice Journal of the American Gastroenterological Association 4, no. 8 (2006): 1035–38, https://doi.org/10.1016/j.cgh.2006.03.020.↩︎
Passildas Jahanmohan Judith et al., “Curcumin’s Effect in Advanced and Metastatic Breast Cancer Patients Treated with First or Second-Line Docetaxel: A Randomized Trial,” Health Science Reports 7, no. 9 (2024): e70052, https://doi.org/10.1002/hsr2.70052.↩︎
Chiara de Waure et al., “Exploring the Contribution of Curcumin to Cancer Therapy: A Systematic Review of Randomized Controlled Trials,” Pharmaceutics 15, no. 4 (2023): 1275, https://doi.org/10.3390/pharmaceutics15041275.↩︎
yair lotan, A Randomized, Double-Blind, Placebo-Controlled Trial of Curcumin to Prevent Progression of Biopsy Proven, Low-Risk Localized Prostate Cancer Patients Undergoing Active Surveillance, Clinical trial registration no. NCT03769766, with University of Texas Southwestern Medical Center (clinicaltrials.gov, 2025), https://clinicaltrials.gov/study/NCT03769766.↩︎
Weidong Liu et al., “Oral Bioavailability of Curcumin: Problems and Advancements,” Journal of Drug Targeting 24, no. 8 (2016): 694–702, https://doi.org/10.3109/1061186X.2016.1157883.↩︎
Young-Min Lee and Yoona Kim, “Is Curcumin Intake Really Effective for Chronic Inflammatory Metabolic Disease? A Review of Meta-Analyses of Randomized Controlled Trials,” Nutrients 16, no. 11 (2024): 1728, https://doi.org/10.3390/nu16111728.↩︎
Adrian L. Lopresti et al., “An Investigation into the Effects of a Curcumin Extract (Curcugen®) on Osteoarthritis Pain of the Knee: A Randomised, Double-Blind, Placebo-Controlled Study,” Nutrients 14, no. 1 (2022): 41, https://doi.org/10.3390/nu14010041.↩︎
Yunes Panahi et al., “Curcuminoids Modify Lipid Profile in Type 2 Diabetes Mellitus: A Randomized Controlled Trial,” Complementary Therapies in Medicine 33 (August 2017): 1–5, https://doi.org/10.1016/j.ctim.2017.05.006.↩︎
Marcia Cruz–Correa et al., “Combination Treatment With Curcumin and Quercetin of Adenomas in Familial Adenomatous Polyposis,” Clinical Gastroenterology and Hepatology 4, no. 8 (2006): 1035–38, https://doi.org/10.1016/j.cgh.2006.03.020.↩︎
Lynne M Howells et al., “Curcumin Combined with FOLFOX Chemotherapy Is Safe and Tolerable in Patients with Metastatic Colorectal Cancer in a Randomized Phase IIa Trial,” The Journal of Nutrition 149, no. 7 (2019): 1133–39, https://doi.org/10.1093/jn/nxz029.↩︎
Reema F. Tayyem et al., “Curcumin Content of Turmeric and Curry Powders,” Nutrition and Cancer 55, no. 2 (2006): 126–31, https://doi.org/10.1207/s15327914nc5502_2.↩︎
“Curcumin and Quercetin Modify Warfarin-Induced Regulation of Porcine CYP1A2 and CYP3A Expression and Activity in Vitro: Xenobiotica: Vol 52 , No 5 – Get Access,” accessed August 12, 2025, https://www.tandfonline.com/doi/full/10.1080/00498254.2022.2089932.↩︎
Peiyu Qiu et al., “Overdose Intake of Curcumin Initiates the Unbalanced State of Bodies,” Journal of Agricultural and Food Chemistry 64, no. 13 (2016): 2765–71, https://doi.org/10.1021/acs.jafc.6b00053.↩︎
Aleksandra Zielińska et al., “Properties, Extraction Methods, and Delivery Systems for Curcumin as a Natural Source of Beneficial Health Effects,” Medicina 56, no. 7 (2020): 336, https://doi.org/10.3390/medicina56070336.↩︎
Mohammad Valizadeh Kiamahalleh et al., “High Performance Curcumin Subcritical Water Extraction from Turmeric (Curcuma Longa L.),” Journal of Chromatography B 1022 (June 2016): 191–98, https://doi.org/10.1016/j.jchromb.2016.04.021.↩︎
Marc_Grossman, “Three Ways to Improve Absorption of Turmeric or Curcumin,” Natural Eye Care Blog: News & Research on Vision, February 2, 2016, https://www.naturaleyecare.com/blog/improve-absorption-turmeric-curcumin/.↩︎
“How Much Curcumin Is in My Turmeric Latte?,” Amoda, accessed August 12, 2025, https://amodatea.com/blogs/amoda-tea-blog/how-much-curcumin-is-in-my-turmeric-latte.↩︎
Ziwei Ma et al., “Pharmaceutical Strategies of Improving Oral Systemic Bioavailability of Curcumin for Clinical Application,” Journal of Controlled Release 316 (December 2019): 359–80, https://doi.org/10.1016/j.jconrel.2019.10.053.↩︎
Masashi Kanai et al., “Dose-Escalation and Pharmacokinetic Study of Nanoparticle Curcumin, a Potential Anticancer Agent with Improved Bioavailability, in Healthy Human Volunteers,” Cancer Chemotherapy and Pharmacology 69, no. 1 (2012): 65–70, https://doi.org/10.1007/s00280-011-1673-1.↩︎
Guido Shoba et al., “Influence of Piperine on the Pharmacokinetics of Curcumin in Animals and Human Volunteers,” Planta Medica 64 (January 2007): 353–56, https://doi.org/10.1055/s-2006-957450.↩︎
“Thorne Curcumin Phytosome (Formerly Meriva),” accessed August 5, 2025, https://store.mayoclinic.com/thorne-meriva-500-sf.html.↩︎
John Cuomo et al., “Comparative Absorption of a Standardized Curcuminoid Mixture and Its Lecithin Formulation,” Journal of Natural Products 74, no. 4 (2011): 664–69, https://doi.org/10.1021/np1007262.↩︎
Alexa Kocher et al., “The Oral Bioavailability of Curcuminoids in Healthy Humans Is Markedly Enhanced by Micellar Solubilisation but Not Further Improved by Simultaneous Ingestion of Sesamin, Ferulic Acid, Naringenin and Xanthohumol,” Journal of Functional Foods 14 (April 2015): 183–91, https://doi.org/10.1016/j.jff.2015.01.045.↩︎
Mangala Hegde et al., “Curcumin Formulations for Better Bioavailability: What We Learned from Clinical Trials Thus Far?,” ACS Omega 8, no. 12 (2023): 10713–46, https://doi.org/10.1021/acsomega.2c07326.↩︎
Masashi Kanai et al., “Dose-Escalation and Pharmacokinetic Study of Nanoparticle Curcumin, a Potential Anticancer Agent with Improved Bioavailability, in Healthy Human Volunteers,” Cancer Chemotherapy and Pharmacology 69, no. 1 (2012): 65–70, https://doi.org/10.1007/s00280-011-1673-1.↩︎
Roodabeh Bahramsoltani et al., “Pharmacokinetic Interactions of Curcuminoids with Conventional Drugs: A Review,” Journal of Ethnopharmacology 209 (September 2017): 1–12, https://doi.org/10.1016/j.jep.2017.07.022.↩︎
“GRAS Notices,” accessed August 25, 2025, https://www.hfpappexternal.fda.gov/scripts/fdcc/index.cfm?id=686&set=GRASNotices&utm_source=chatgpt.com.↩︎
PDQ Integrative, Alternative, and Complementary Therapies Editorial Board, “Curcumin (Curcuma, Turmeric) and Cancer (PDQ®): Patient Version,” in PDQ Cancer Information Summaries (National Cancer Institute (US), 2002), http://www.ncbi.nlm.nih.gov/books/NBK578436/.↩︎
“Office of Dietary Supplements – Omega-3 Fatty Acids,” accessed August 28, 2025, https://ods.od.nih.gov/factsheets/Omega3FattyAcids-HealthProfessional/.↩︎
Sujatha Rajaram, “Health Benefits of Plant-Derived α-Linolenic Acid,” The American Journal of Clinical Nutrition 100, no. suppl_1 (2014): 443S-448S, https://doi.org/10.3945/ajcn.113.071514.↩︎
Jenette Restivo, “Omega-3 Foods: Incorporating Healthy Fats into Your Diet,” Harvard Health, June 14, 2024, https://www.health.harvard.edu/nutrition/omega-3-foods-incorporating-healthy-fats-into-your-diet.↩︎
“Quantitation of Alpha-Linolenic Acid Elongation to Eicosapentaenoic and Docosahexaenoic Acid as Affected by the Ratio of N6/N3 Fatty Acids | Nutrition & Metabolism | Full Text,” accessed August 28, 2025, https://nutritionandmetabolism.biomedcentral.com/articles/10.1186/1743-7075-6-8.↩︎
Rebeca Caires et al., “Omega-3 Fatty Acids Modulate TRPV4 Function through Plasma Membrane Remodeling,” Cell Reports 21, no. 1 (2017): 246–58, https://doi.org/10.1016/j.celrep.2017.09.029.↩︎
Philip C. Calder, “Omega-3 Fatty Acids and Inflammatory Processes: From Molecules to Man,” Biochemical Society Transactions 45, no. 5 (2017): 1105–15, https://doi.org/10.1042/BST20160474.↩︎
WebMD Editorial Contributor, “Healthy Foods High in Omega-6,” WebMD, accessed October 22, 2025, https://www.webmd.com/diet/foods-high-in-omega-6.↩︎
E. Patterson et al., “Health Implications of High Dietary Omega-6 Polyunsaturated Fatty Acids,” Journal of Nutrition and Metabolism 2012 (2012): 539426, https://doi.org/10.1155/2012/539426.↩︎
A. P Simopoulos, “The Importance of the Ratio of Omega-6/Omega-3 Essential Fatty Acids,” Biomedicine & Pharmacotherapy 56, no. 8 (2002): 365–79, https://doi.org/10.1016/S0753-3322(02)00253-6.↩︎
Ivana Djuricic and Philip C. Calder, “Beneficial Outcomes of Omega-6 and Omega-3 Polyunsaturated Fatty Acids on Human Health: An Update for 2021,” Nutrients 13, no. 7 (2021): 2421, https://doi.org/10.3390/nu13072421.↩︎
Hee-Yong Kim et al., “Molecular and Signaling Mechanisms for Docosahexaenoic Acid-Derived Neurodevelopment and Neuroprotection,” International Journal of Molecular Sciences 23, no. 9 (2022): 4635, https://doi.org/10.3390/ijms23094635.↩︎
R. Preston Mason et al., “Emerging Mechanisms of Cardiovascular Protection for the Omega-3 Fatty Acid Eicosapentaenoic Acid,” Arteriosclerosis, Thrombosis, and Vascular Biology 40, no. 5 (2020): 1135–47, https://doi.org/10.1161/ATVBAHA.119.313286.↩︎
Ahmed A. Daak et al., “Omega 3 (n − 3) Fatty Acids down-Regulate Nuclear Factor-Kappa B (NF-κB) Gene and Blood Cell Adhesion Molecule Expression in Patients with Homozygous Sickle Cell Disease,” Blood Cells, Molecules, and Diseases 55, no. 1 (2015): 48–55, https://doi.org/10.1016/j.bcmd.2015.03.014.↩︎
“NIH-AARP Diet and Health Study – NCI,” cgvArticle, November 18, 2022, nciglobal,ncienterprise, https://dceg.cancer.gov/research/who-we-study/nih-aarp-diet-health-study.↩︎
Claire Bosire et al., “Index-Based Dietary Patterns and the Ribroad framework, fish consumption and long-chain omega-3 fatty acid intake were extensively studied as potential risk of Prostate Cancer in the NIH-AARP Diet and Health Study,” American Journal of Epidemiology 177, no. 6 (2013): 504–13, https://doi.org/10.1093/aje/kws261.↩︎
“Singapore Chinese Health Study (SCHS),” Saw Swee Hock School of Public Health, n.d., accessed September 22, 2025, https://sph.nus.edu.sg/research/cohort-schs/.↩︎
M. Gago-Dominguez et al., “Opposing Effects of Dietary N-3 and n-6 Fatty Acids on Mammary Carcinogenesis: The Singapore Chinese Health Study,” British Journal of Cancer 89, no. 9 (2003): 1686–92, https://doi.org/10.1038/sj.bjc.6601340.↩︎
Emily White et al., “VITamins And Lifestyle Cohort Study: Study Design and Characteristics of Supplement Users,” American Journal of Epidemiology 159, no. 1 (2004): 83–93, https://doi.org/10.1093/aje/kwh010.↩︎
Theodore M. Brasky et al., “Specialty Supplements and Breast Cancer Risk in the VITamins And Lifestyle (VITAL) Cohort,” Cancer Epidemiology, Biomarkers & Prevention: A Publication of the American Association for Cancer Research, Cosponsored by the American Society of Preventive Oncology 19, no. 7 (2010): 1696–708, https://doi.org/10.1158/1055-9965.EPI-10-0318.↩︎
Elizabeth D. Kantor et al., “Long-Chain Omega-3 Polyunsaturated Fatty Acid Intake and Risk of Colorectal Cancer,” Nutrition and Cancer 66, no. 4 (2014): 716–27, https://doi.org/10.1080/01635581.2013.804101.↩︎
JoAnn E. Manson et al., “Principal Results of the VITamin D and OmegA-3 TriaL (VITAL) and Updated Meta-Analyses of Relevant Vitamin D Trials,” The Journal of Steroid Biochemistry and Molecular Biology 198 (April 2020): 105522, https://doi.org/10.1016/j.jsbmb.2019.105522.↩︎
William J. Aronson et al., “High Omega-3, Low Omega-6 Diet With Fish Oil for Men With Prostate Cancer on Active Surveillance: The CAPFISH-3 Randomized Clinical Trial,” Journal of Clinical Oncology: Official Journal of the American Society of Clinical Oncology 43, no. 7 (2025): 800–809, https://doi.org/10.1200/JCO.24.00608.↩︎
University of Kansas Medical Center, Omega 3 Fatty Acids in Colorectal Cancer (CRC) Prevention in Patients With Lynch Syndrome (COLYNE), Clinical trial registration no. NCT03831698 (clinicaltrials.gov, 2023), https://clinicaltrials.gov/study/NCT03831698.↩︎
“Prostate Cancer Prevention Trial (PCPT): Questions and Answers – NCI,” accessed September 22, 2025, https://www.cancer.gov/types/prostate/research/prostate-cancer-prevention-trial-qa.↩︎
Theodore M. Brasky et al., “Plasma Phospholipid Fatty Acids and Prostate Cancer Risk in the SELECT Trial,” Journal of the National Cancer Institute 105, no. 15 (2013): 1132–41, https://doi.org/10.1093/jnci/djt174.↩︎
“Office of Dietary Supplements – Omega-3 Fatty Acids,” accessed October 6, 2025, https://ods.od.nih.gov/factsheets/Omega3FattyAcids-HealthProfessional/.↩︎
Organisation des Nations Unies pour l’alimentation et l’agriculture and Organisation mondiale de la santé, eds., Fats and Fatty Acids in Human Nutrition: Report of an Expert Consultation 10-14 November 2008, Geneva, FAO Food and Nutrition Paper 91 (FAO, 2010).↩︎
EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA), “Scientific Opinion on the Tolerable Upper Intake Level of Eicosapentaenoic Acid (EPA), Docosahexaenoic Acid (DHA) and Docosapentaenoic Acid (DPA),” EFSA Journal 10, no. 7 (2012): 2815, https://doi.org/10.2903/j.efsa.2012.2815.↩︎
Penny M. Kris-Etherton et al., “Fish Consumption, Fish Oil, Omega-3 Fatty Acids, and Cardiovascular Disease,” Circulation 106, no. 21 (2002): 2747–57, https://doi.org/10.1161/01.CIR.0000038493.65177.94.↩︎
K C H Fearon et al., “Effect of a Protein and Energy Dense N-3 Fatty Acid Enriched Oral Supplement on Loss of Weight and Lean Tissue in Cancer Cachexia: A Randomised Double Blind Trial,” Gut 52, no. 10 (2003): 1479–86, https://doi.org/10.1136/gut.52.10.1479.↩︎
Rachel A. Murphy et al., “Supplementation with Fish Oil Increases First-Line Chemotherapy Efficacy in Patients with Advanced Nonsmall Cell Lung Cancer,” Cancer 117, no. 16 (2011): 3774–80, https://doi.org/10.1002/cncr.25933.↩︎
Carolina de Quadros Camargo et al., “Fish Oil Supplementation during Chemotherapy Increases Posterior Time to Tumor Progression in Colorectal Cancer,” Nutrition and Cancer 68, no. 1 (2016): 70–76, https://doi.org/10.1080/01635581.2016.1115097.↩︎
Michel Carlos Mocellin et al., “Fish Oil Decreases C-Reactive Protein/Albumin Ratio Improving Nutritional Prognosis and Plasma Fatty Acid Profile in Colorectal Cancer Patients,” Lipids 48, no. 9 (2013): 879–88, https://doi.org/10.1007/s11745-013-3816-0.↩︎
Jann Arends et al., “ESPEN Guidelines on Nutrition in Cancer Patients,” Clinical Nutrition 36, no. 1 (2017): 11–48, https://doi.org/10.1016/j.clnu.2016.07.015.↩︎
Maurizio Muscaritoli et al., “ESPEN Practical Guideline: Clinical Nutrition in Cancer,” Clinical Nutrition 40, no. 5 (2021): 2898–913, https://doi.org/10.1016/j.clnu.2021.02.005.↩︎
Breanne M Anderson and David WL Ma, “Are All N-3 Polyunsaturated Fatty Acids Created Equal?,” Lipids in Health and Disease 8 (August 2009): 33, https://doi.org/10.1186/1476-511X-8-33.↩︎
“Tolerable Upper Intake Level of EPA, DHA and DPA | EFSA,” July 27, 2012, https://www.efsa.europa.eu/en/efsajournal/pub/2815.↩︎
Deepak L. Bhatt et al., “Cardiovascular Risk Reduction with Icosapent Ethyl for Hypertriglyceridemia,” New England Journal of Medicine 380, no. 1 (2019): 11–22, https://doi.org/10.1056/NEJMoa1812792.↩︎
William S. Harris and Clemens Von Schacky, “The Omega-3 Index: A New Risk Factor for Death from Coronary Heart Disease?,” Preventive Medicine 39, no. 1 (2004): 212–20, https://doi.org/10.1016/j.ypmed.2004.02.030.↩︎
Roxanne Karimi et al., “A Quantitative Synthesis of Mercury in Commercial Seafood and Implications for Exposure in the United States,” Environmental Health Perspectives 120, no. 11 (2012): 1512–19, https://doi.org/10.1289/ehp.1205122.↩︎
Organisation des Nations Unies pour l’alimentation et l’agriculture and Organisation mondiale de la santé, eds., Fats and Fatty Acids in Human Nutrition: Report of an Expert Consultation 10-14 November 2008, Geneva, FAO Food and Nutrition Paper 91 (FAO, 2010).↩︎
Zlata Kralik et al., “Natural Multi-Enriched Eggs with n-3 Polyunsaturated Fatty Acids, Selenium, Vitamin E, and Lutein,” Animals 13, no. 2 (2023), https://doi.org/10.3390/ani13020321.https://doi.org/10.3390/ani13020321.s↩︎
Lam Phuoc Thanh et al., “Effect of Fish Oil and Linseed Oil on Intake, Milk Yield and Milk Fatty Acid Profile in Goats,” Animals : An Open Access Journal from MDPI 13, no. 13 (2023): 2174, https://doi.org/10.3390/ani13132174.↩︎
Ankit Goyal et al., “Fortification of Dahi (Indian Yoghurt) with Omega-3 Fatty Acids Using Microencapsulated Flaxseed Oil Microcapsules,” Journal of Food Science and Technology 53, no. 5 (2016): 2422–33, https://doi.org/10.1007/s13197-016-2220-1.↩︎
Ashish Pandey et al., “Omega-3 Fatty Acid Fortification of Plant-Based Beverages to Enhance Their Nutritional Profile,” Foods 14, no. 9 (2025): 1602, https://doi.org/10.3390/foods14091602.↩︎
Human Foods Program, “Advice about Eating Fish,” FDA, FDA, September 5, 2024, https://www.fda.gov/food/consumers/advice-about-eating-fish.↩︎
Laurie Chevalier et al., “Pharmacokinetics of Supplemental Omega-3 Fatty Acids Esterified in Monoglycerides, Ethyl Esters, or Triglycerides in Adults in a Randomized Crossover Trial,” The Journal of Nutrition 151, no. 5 (2021): 1111–18, https://doi.org/10.1093/jn/nxaa458.↩︎
Jan Philipp Schuchardt et al., “Incorporation of EPA and DHA into Plasma Phospholipids in Response to Different Omega-3 Fatty Acid Formulations – a Comparative Bioavailability Study of Fish Oil vs. Krill Oil,” Lipids in Health and Disease 10, no. 1 (2011): 145, https://doi.org/10.1186/1476-511X-10-145.↩︎
Anton Köhler et al., “Bioavailability of Fatty Acids from Krill Oil, Krill Meal and Fish Oil in Healthy Subjects–a Randomized, Single-Dose, Cross-over Trial,” Lipids in Health and Disease 14, no. 1 (2015): 19, https://doi.org/10.1186/s12944-015-0015-4.↩︎
Linda M. Arterburn et al., “Bioequivalence of Docosahexaenoic Acid from Different Algal Oils in Capsules and in a DHA-Fortified Food,” Lipids 42, no. 11 (2007): 1011, https://doi.org/10.1007/s11745-007-3098-5.↩︎
“FISH OIL: Overview, Uses, Side Effects, Precautions, Interactions, Dosing and Reviews,” accessed October 20, 2025, https://www.webmd.com/vitamins/ai/ingredientmono-993/fish-oil.↩︎
Kristina N. Krupa et al., “Omega-3 Fatty Acids,” in StatPearls (StatPearls Publishing, 2025), http://www.ncbi.nlm.nih.gov/books/NBK564314/.↩︎
Mustafa Javaid et al., “Bleeding Risk in Patients Receiving Omega‐3 Polyunsaturated Fatty Acids: A Systematic Review and Meta‐Analysis of Randomized Clinical Trials,” Journal of the American Heart Association 13, no. 10 (2024): e032390, https://doi.org/10.1161/JAHA.123.032390.↩︎
Baris Gencer et al., “Effect of Long-Term Marine Omega-3 Fatty Acids Supplementation on the Risk of Atrial Fibrillation in Randomized Controlled Trials of Cardiovascular Outcomes: A Systematic Review and Meta-Analysis,” Circulation 144, no. 25 (2021): 1981–90, https://doi.org/10.1161/CIRCULATIONAHA.121.055654.↩︎
Ann C. Skulas-Ray et al., “Omega-3 Fatty Acids for the Management of Hypertriglyceridemia: A Science Advisory From the American Heart Association,” Circulation 140, no. 12 (2019): e673–91, https://doi.org/10.1161/CIR.0000000000000709.↩︎
“Excess Omega-3 Fatty Acids Could Lead to Negative Health Effects | Newsroom | Oregon State University,” October 28, 2013, https://news.oregonstate.edu/news/excess-omega-3-fatty-acids-could-lead-negative-health-effects.↩︎
“Vitamin A,” in LiverTox: Clinical and Research Information on Drug-Induced Liver Injury (National Institute of Diabetes and Digestive and Kidney Diseases, 2012), http://www.ncbi.nlm.nih.gov/books/NBK548165/.↩︎
“GRAS Notices,” accessed October 23, 2025, https://hfpappexternal.fda.gov/scripts/fdcc/index.cfm?id=200&set=GRASNotices&utm_source=chatgpt.com.↩︎
“Omega-3 Supplements: What You Need To Know,” NCCIH, accessed October 23, 2025, https://www.nccih.nih.gov/health/omega3-supplements-what-you-need-to-know.↩︎
“Drug Approval Package,” accessed October 23, 2025, https://www.accessdata.fda.gov/drugsatfda_docs/nda/2012/202057Orig1s000TOC.cfm.↩︎
“Definition of Sulforaphane – NCI Drug Dictionary – NCI,” nciAppModulePage, February 2, 2011, nciglobal,ncienterprise, https://www.cancer.gov/publications/dictionaries/cancer-drug/def/sulforaphane.↩︎
Jed W. Fahey et al., “Broccoli Sprouts: An Exceptionally Rich Source of Inducers of Enzymes That Protect against Chemical Carcinogens,” Proceedings of the National Academy of Sciences 94, no. 19 (1997): 10367–72, https://doi.org/10.1073/pnas.94.19.10367.↩︎
Jae Kwang Kim and Sang Un Park, “Current Potential Health Benefits of Sulforaphane,” EXCLI Journal 15 (October 2016): 571–77, https://doi.org/10.17179/excli2016-485.↩︎
Y. Zhang et al., “A Major Inducer of Anticarcinogenic Protective Enzymes from Broccoli: Isolation and Elucidation of Structure,” Proceedings of the National Academy of Sciences of the United States of America 89, no. 6 (1992): 2399–403, https://doi.org/10.1073/pnas.89.6.2399.↩︎
Christine A. Houghton, “Sulforaphane: Its ‘Coming of Age’ as a Clinically Relevant Nutraceutical in the Prevention and Treatment of Chronic Disease,” Oxidative Medicine and Cellular Longevity 2019 (October 2019): 2716870, https://doi.org/10.1155/2019/2716870.↩︎
Tetiana R. Dmytriv et al., “Glucoraphanin Conversion into Sulforaphane and Related Compounds by Gut Microbiota,” Frontiers in Physiology 16 (February 2025), https://doi.org/10.3389/fphys.2025.1497566.↩︎
Chenqi Hu et al., “Modification of Keap1 Cysteine Residues by Sulforaphane,” Chemical Research in Toxicology 24, no. 4 (2011): 515–21, https://doi.org/10.1021/tx100389r.↩︎
Albena T. Dinkova-Kostova et al., “Direct Evidence That Sulfhydryl Groups of Keap1 Are the Sensors Regulating Induction of Phase 2 Enzymes That Protect against Carcinogens and Oxidants,” Proceedings of the National Academy of Sciences of the United States of America 99, no. 18 (2002): 11908–13, https://doi.org/10.1073/pnas.172398899.↩︎
Elke Heiss et al., “Nuclear Factor κB Is a Molecular Target for Sulforaphane-Mediated Anti-Inflammatory Mechanisms *,” Journal of Biological Chemistry 276, no. 34 (2001): 32008–15, https://doi.org/10.1074/jbc.M104794200.↩︎
Melinda C. Myzak et al., “A Novel Mechanism of Chemoprotection by Sulforaphane: Inhibition of Histone Deacetylase,” Cancer Research 64, no. 16 (2004): 5767–74, https://doi.org/10.1158/0008-5472.CAN-04-1326.↩︎
Ajita V. Singh et al., “Sulforaphane Induces Caspase-Mediated Apoptosis in Cultured PC-3 Human Prostate Cancer Cells and Retards Growth of PC-3 Xenografts in Vivo,” Carcinogenesis 25, no. 1 (2004): 83–90, https://doi.org/10.1093/carcin/bgg178.↩︎
Muhammad Asif Ali et al., “Anticancer Properties of Sulforaphane: Current Insights at the Molecular Level,” Frontiers in Oncology 13 (June 2023), https://doi.org/10.3389/fonc.2023.1168321.↩︎
Christine A. Houghton, “The Rationale for Sulforaphane Favourably Influencing Gut Homeostasis and Gut–Organ Dysfunction: A Clinician’s Hypothesis,” International Journal of Molecular Sciences 24, no. 17 (2023): 13448, https://doi.org/10.3390/ijms241713448.↩︎
Egner Pa et al., “Rapid and Sustainable Detoxication of Airborne Pollutants by Broccoli Sprout Beverage: Results of a Randomized Clinical Trial in China,” Cancer Prevention Research (Philadelphia, Pa.) 7, no. 8 (2014), https://doi.org/10.1158/1940-6207.CAPR-14-0103.↩︎
Akinori Yanaka et al., “Dietary Sulforaphane-Rich Broccoli Sprouts Reduce Colonization and Attenuate Gastritis in Helicobacter Pylori–Infected Mice and Humans,” Cancer Prevention Research 2, no. 4 (2009): 353–60, https://doi.org/10.1158/1940-6207.CAPR-08-0192.↩︎
Lauren L. Atwell et al., “Sulforaphane Bioavailability and Chemopreventive Activity in Women Scheduled for Breast Biopsy,” Cancer Prevention Research (Philadelphia, Pa.) 8, no. 12 (2015): 1184–91, https://doi.org/10.1158/1940-6207.CAPR-15-0119.↩︎
Joshi J. Alumkal et al., “A Phase II Study of Sulforaphane-Rich Broccoli Sprout Extracts in Men with Recurrent Prostate Cancer,” Investigational New Drugs 33, no. 2 (2015): 480–89, https://doi.org/10.1007/s10637-014-0189-z.↩︎
Bernard G. Cipolla et al., “Effect of Sulforaphane in Men with Biochemical Recurrence after Radical Prostatectomy,” Cancer Prevention Research 8, no. 8 (2015): 712–19, https://doi.org/10.1158/1940-6207.CAPR-14-0459.↩︎
Christine A. Houghton, “The Rationale for Sulforaphane Favourably Influencing Gut Homeostasis and Gut-Organ Dysfunction: A Clinician’s Hypothesis,” International Journal of Molecular Sciences 24, no. 17 (2023): 13448, https://doi.org/10.3390/ijms241713448.↩︎
Jian-Min Yuan et al., “Randomized Phase II Clinical Trial of Sulforaphane in Former Smokers at High Risk for Lung Cancer,” Cancer Prevention Research (Philadelphia, Pa.) 18, no. 6 (2025): 335–45, https://doi.org/10.1158/1940-6207.CAPR-24-0386.↩︎
H. Lee Moffitt Cancer Center and Research Institute, Randomized, Phase II Clinical Trial of Sulforaphane in Bladder Cancer Chemoprevention, Clinical trial registration no. NCT03517995, with National Institutes of Health (NIH) and Johns Hopkins University (clinicaltrials.gov, 2020), https://clinicaltrials.gov/study/NCT03517995.↩︎
“Testing the Effect of Sulforaphane, a Compound Naturally Found in Cruciferous Vegetables, on Preventing Melanoma in Patients With a Prior History of Melanoma,” accessed November 13, 2025, https://prevention.cancer.gov/clinical-trials/clinical-trials-search/nct07040280.↩︎
John D. Clarke et al., “Multi-Targeted Prevention of Cancer by Sulforaphane,” Cancer Letters, Natural Products Special Issue, vol. 269, no. 2 (2008): 291–304, https://doi.org/10.1016/j.canlet.2008.04.018.↩︎
Pa et al., “Rapid and Sustainable Detoxication of Airborne Pollutants by Broccoli Sprout Beverage.”↩︎
Akinori Yanaka et al., “Dietary Sulforaphane-Rich Broccoli Sprouts Reduce Colonization and Attenuate Gastritis in Helicobacter Pylori-Infected Mice and Humans,” Cancer Prevention Research (Philadelphia, Pa.) 2, no. 4 (2009): 353–60, https://doi.org/10.1158/1940-6207.CAPR-08-0192.↩︎
Kim and Park, “Current Potential Health Benefits of Sulforaphane.”↩︎
Alumkal et al., “A Phase II Study of Sulforaphane-Rich Broccoli Sprout Extracts in Men with Recurrent Prostate Cancer.”↩︎
Yoko Yagishita et al., “Broccoli or Sulforaphane: Is It the Source or Dose That Matters?,” Molecules 24, no. 19 (2019): 3593, https://doi.org/10.3390/molecules24193593.↩︎
Dana ElKhalifa et al., “Efficacy and Tolerability of Sulforaphane in the Therapeutic Management of Cancers: A Systematic Review of Randomized Controlled Trials,” Frontiers in Oncology 13 (November 2023), https://doi.org/10.3389/fonc.2023.1251895.↩︎
Nieves Baenas et al., “Influence of Cooking Methods on Glucosinolates and Isothiocyanates Content in Novel Cruciferous Foods,” Foods 8, no. 7 (2019): 257, https://doi.org/10.3390/foods8070257.↩︎
Dushani L. Palliyaguru et al., “Isothiocyanates: Translating the Power of Plants to People,” Molecular Nutrition & Food Research 62, no. 18 (2018): 1700965, https://doi.org/10.1002/mnfr.201700965.↩︎
“Sulforaphane: Benefits, Side Effects, and Food Sources,” Healthline, February 26, 2019, https://www.healthline.com/nutrition/sulforaphane.↩︎
J. W. Fahey et al., “Broccoli Sprouts: An Exceptionally Rich Source of Inducers of Enzymes That Protect against Chemical Carcinogens,” Proceedings of the National Academy of Sciences of the United States of America 94, no. 19 (1997): 10367–72, https://doi.org/10.1073/pnas.94.19.10367.↩︎
Omobolanle O. Oloyede et al., “The Impact of Domestic Cooking Methods on Myrosinase Stability, Glucosinolates and Their Hydrolysis Products in Different Cabbage (Brassica Oleracea) Accessions,” Foods 10, no. 12 (2021): 2908, https://doi.org/10.3390/foods10122908.↩︎
“Gut Myrosinases Convert Glucosinolates to Sulforaphane.,” FoundMyFitness, March 22, 2022, https://www.foundmyfitness.com/stories/qtxhrl.↩︎
Sameer Khalil Ghawi et al., “The Potential to Intensify Sulforaphane Formation in Cooked Broccoli (Brassica Oleracea Var. Italica) Using Mustard Seeds (Sinapis Alba),” Food Chemistry 138, no. 2 (2013): 1734–41, https://doi.org/10.1016/j.foodchem.2012.10.119.↩︎
Duygu Ağagündüz et al., “Cruciferous Vegetables and Their Bioactive Metabolites: From Prevention to Novel Therapies of Colorectal Cancer,” Evidence-Based Complementary and Alternative Medicine 2022, no. 1 (2022): 1534083, https://doi.org/10.1155/2022/1534083.↩︎
John A Bouranis et al., “Sulforaphane and Sulforaphane-Nitrile Metabolism in Humans Following Broccoli Sprout Consumption: Inter-Individual Variation, Association with Gut Microbiome Composition, and Differential Bioactivity,” Molecular Nutrition & Food Research 68, no. 4 (2024): e2300286, https://doi.org/10.1002/mnfr.202300286.↩︎
A Shapiro et al., Human Metabolism and Excretion of Cancer Chemoprotective Glucosinolates and Isothiocyanates of Cruciferous, n.d.↩︎
“Prostaphane,” Prostaphane, n.d., accessed November 17, 2025, https://www.prostaphane.com/prostaphane/.↩︎
Cipolla et al., “Effect of Sulforaphane in Men with Biochemical Recurrence after Radical Prostatectomy.”↩︎
Wu Yuanfeng et al., “Approaches for Enhancing the Stability and Formation of Sulforaphane,” Food Chemistry 345 (May 2021): 128771, https://doi.org/10.1016/j.foodchem.2020.128771.↩︎
www.prohealth.com, “Liposomal Sulforaphane: A Broccoli-Based Antioxidant with Enhanced Absorption That Supports Health With Age,” ProHealth.Com, accessed November 17, 2025, https://www.prohealth.com/blogs/control-how-you-age/liposomal-sulforaphane-a-broccoli-based-antioxidant-with-enhanced-absorption-that-supports-health-with-age.↩︎
“Sulforaphane – an Overview | ScienceDirect Topics,” accessed November 17, 2025, https://www.sciencedirect.com/topics/pharmacology-toxicology-and-pharmaceutical-science/sulforaphane.↩︎
Masahiko Ishida et al., “Glucosinolate Metabolism, Functionality and Breeding for the Improvement of Brassicaceae Vegetables,” Breeding Science 64, no. 1 (May 2014): 48–59, https://doi.org/10.1270/jsbbs.64.48.↩︎
Katarzyna Lubelska et al., “Interactions between Drugs and Sulforaphane Modulate the Drug Metabolism Enzymatic System,” Pharmacological Reports 64, no. 5 (2012): 1243–52, https://doi.org/10.1016/S1734-1140(12)70920-9.↩︎
Paweł Paśko et al., “Broccoli Sprouts and Their Influence on Thyroid Function in Different In Vitro and In Vivo Models,” Plants 11, no. 20 (2022): 2750, https://doi.org/10.3390/plants11202750.ng↩︎
L. Schwingshackl and G. Hoffmann, “Mediterranean Dietary Pattern, Inflammation and Endothelial Function: A Systematic Review and Meta-Analysis of Intervention Trials,” Nutrition, Metabolism and Cardiovascular Diseases 24, no. 9 (September 2014): 929–39, https://doi.org/10.1016/j.numecd.2014.03.003.↩︎
Ramón Estruch et al., “Retraction and Republication: Primary Prevention of Cardiovascular Disease with a Mediterranean Diet. N Engl J Med 2013;368:1279-90.,” New England Journal of Medicine 378, no. 25 (June 2018): 2441–42, https://doi.org/10.1056/NEJMc1806491.↩︎
Giuseppe Grosso et al., “Mediterranean Diet and Cancer: Epidemiological Evidence and Mechanism of Selected Aspects,” BMC Surgery 13 Suppl 2, no. Suppl 2 (2013): S14, https://doi.org/10.1186/1471-2482-13-S2-S14.↩︎
Philip C. Calder, “Dietary Factors and Low-Grade Inflammation in Relation to Overweight and Obesity Revisited,” The British Journal of Nutrition 127, no. 10 (n.d.): 1455–57, https://doi.org/10.1017/S0007114522000782.↩︎
Gauree G. Konijeti et al., “Efficacy of the Autoimmune Protocol Diet for Inflammatory Bowel Disease,” Inflammatory Bowel Diseases 23, no. 11 (November 2017): 2054–60, https://doi.org/10.1097/MIB.0000000000001221.↩︎
